biologista

Sexe : 
 Messages : 180 Messages : 180
 Points : 317 Points : 317
 Date de naissance : 07/05/1992 Date de naissance : 07/05/1992
 Age : 32 Age : 32
 Emploi : Chez mon papa Emploi : Chez mon papa
 | Sujet: La cellule végétale  Jeu 6 Déc 2012 - 0:26 Jeu 6 Déc 2012 - 0:26 | |
| souce : biodis I. INTRODUCTION-RAPPEL 1. Procaryotes/EucaryotesLes
premiers êtres vivants, apparus sur terre il y a environ 3,5 milliards
d'années, étaient des bactéries dépourvus de noyau cellulaire : des
procaryotes. C'est à partir de ces procaryotes que des êtres plus
complexes sont apparus : les eucaryotes. Leur caractéristique
essentielle est de posséder un noyau cellulaire qui renferme l'ADN,
support de l'information génétique.Tous les animaux et les végétaux sont
des eucaryotes. Les principaux caractères distinctifs entre eucaryotes et procaryotes sont résumés dans le tableau suivant :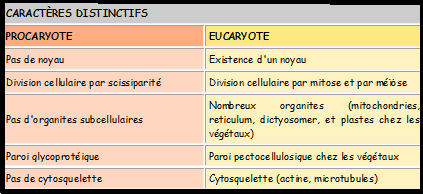 Tableau 1 Tableau 1
discipline biologique, dont fait partie la botanique, a pour objet
d’étude des êtres vivants. Le monde vivant se différencie du monde
minéral par un ensemble de caractéristiques importantes.Ce monde vivant est habituellement ( WITTHAKER ) divisé en cinq règnes : LE REGNE DES MONERES
Tous les organismes procaryotes, à savoir les bactéries et les
cyanobactéries ( autrefois appelées algues bleues), font partie du règne
des Monères. Les procaryotes n’ont pas de membrane nucléaire, ni de
mitochondries ni d’autres organites membranaires présents dans les
cellules eucaryotes.Leur ADN forme un double hélice circulaire
pratiquement dénuée de protéines . Les monères se divisent et se
reproduisent par fission ( ou scissiparité ), sans présenter les
divisions nucléaires mitotiques ou méiotiques propres aux eucaryotes.
Les procaryotes se distinguent aussi bio-chimiquement des eucaryotes,
par les matériaux formant les parois cellulaires, par la taille et la
composition de leurs ribosomes, ainsi que par certaines voies
métaboliques.LE REGNE DES PROTISTES
Les protistes sont des organismes unicellulaires eucaryotes ; ils ont
donc une membrane nucléaire et des chromosomes linéaires ( composés
d’ADN et de protéines). Ils peuvent se diviser par mitose et, pour la
plupart, par méiose . Les premiers eucaryotes étaient sans aucun doute
des protistes ; certains sont à l’origine des mycètes, des végétaux et
des animaux, qui représentent actuellement les formes de vie dominantes
de la terre.Les protistes actuels ont des modes de vie assez semblables à
ceux des eucaryotes supérieurs : certains sont photosynthétiques,
d’autres se nourrissent par ingestion et d’autres encore par
absorption ; certains sont mobiles alors que d’autres ne le sont pas ;
certains ont une paroi cellulaire, d’autres pas ; toutes les
combinaisons de ces caractéristiques se retrouvent chez divers
représentants de ce règne.LE REGNE DES MYCETES
On classe souvent les mycètes avec les végétaux parce qu’ils sont
immobiles et ont une paroi externe qui ressemble à bien des égard à la
paroi cellulaire des végétaux. Toutefois, les mycètes ne peuvent
élaborer leurs propres aliments : ils doivent absorber leur nourriture à
partir d’une source organique, vivante ou non. Dans bons nombres de cas
, avant de pouvoir absorber leurs aliments, il sécrètent des enzymes
digestives qui les dégradent à l’extérieur d’eux. Witthaker avait
l’impression qu’il fallait séparer complètement les mycètes des
végétaux, parce qu’ils ne se nourrissent que par absorption , alors que
les végétaux ne se nourrissent par absorption que de manière accessoire
et qu’ils élaborent leurs propres molécules organiques. En outre les
mycètes se distinguent des végétaux sur le plan de la reproduction, de
la structure corporelle et de la composition de leur paroi cellulaire.LE REGNE DES VEGETAUX
Tous les organismes du règne des végétaux sont des eucaryotes pourvus
de parois cellulaires contenant de la cellulose. La plupart ont des
pigments chlorophylliens et font de la photosynthèse dans de
chloroplastes, quelques espèces ont cependant perdu leur chlorophylle et
se nourrissent par absorption, à la façon des mycètes. Ce règne
comprend les algues pluricellulaires ainsi que les plantes terrestres
plus familières – les mousses, les fougères, les graminées, les arbustes
et les arbres.LE REGNE DES ANIMAUX
Les animaux sont des eucaryotes pluricellulaires et hétérotrophes qui
dépendent d’autres organismes comme sources de composés organiques, et
qui se nourrissent principalement paringestion. La plupart des animaux
peuvent se mouvoir. Tous les animaux, à l’exception des plus simples,
produisent des gamètes ( ovules et spermatozoïdes) dans des organes
pluricellulaires et les œufs et les œufs se développent pour former des
embryons pluricellulaires. 3. La biologie végétaleTraditionnellement,la
biologie végétale traite de l’étude des végétaux et parfois des
champignons par l’intermédiaire de la mycologie.La biologie comporte des
disciplines fondamentales dont les bases sont communes aux différents
règnes ( Chimie biologique, biologie moléculaire, génétique,biologie
cellulaire ), mais également des branches spécifiques a ces différents règnes, ainsi, on parle de : Biologie végétale et de biologie animale .Biologie végétale :Quoique
généralement semblables, les cellules végétales et animales se
différencient par différents points que nous avons examinés en biologie
cellulaire, si bien que l’on doit concevoir une cytologie particulière
pour les végétaux, et également une histologie végétale, génétique
végétale, écologie végétale, physiologie végétale …etc II. LA CYTOLOGIE VEGETALE II.1 La cellule, aspect structurelII.1.1 Base Noyau,
cytoplasme, paroi cellulaire ; nombreux organites cytoplasmiques ou
inclusions : réticulum endoplasmique, appareil de Golgi,
mitochondries,lysosomes, inclusions huileuses, vacuoles, plastes. La cellule végétale se distingue de la cellule animale par quatre caractéristiques cytologiques majeures ( voir biologie cellulaire) :la vacuole fortement développée les plastes et pigments assimilateurs (chlorophylles, caroténoïdes,phycobilines) la paroi, mitoyenne, épaisse et rigide ( cellulose).Absence de centrosomes et appareil de Golgi moins apparent.II.1.2 Schémas d’une cellule végétale théorique ( avec ses caractéristiques générales les plus importantes).II.1.2.1 La cellule jeune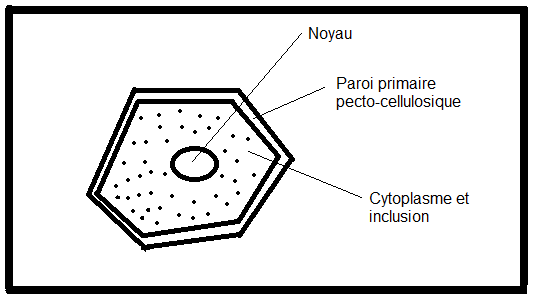 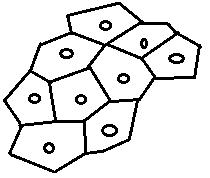 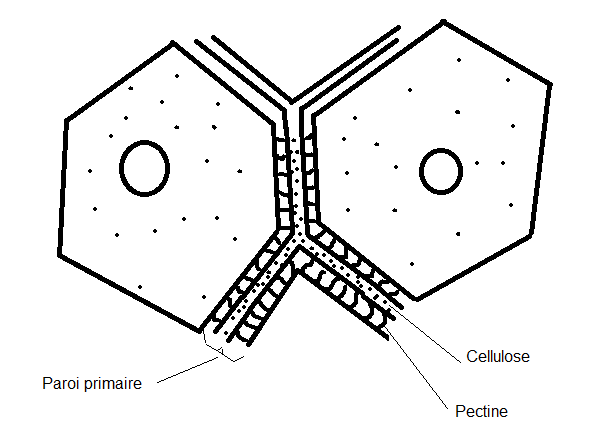 Sur le schéma représentant la membrane primaire, on remarque que l’accolement de plusieurs cellules fait apparaître une structure constituée de trois couches : cellulose-pectine-cellulose , c’est la paroi primaire ou pectocellulosique, celle-ci est constituée de cellulose, de pectines et d’hémicellulose ( voir B. cellulaire). II.1.2.2 Composition de paroiPECTINES :sont Sur le schéma représentant la membrane primaire, on remarque que l’accolement de plusieurs cellules fait apparaître une structure constituée de trois couches : cellulose-pectine-cellulose , c’est la paroi primaire ou pectocellulosique, celle-ci est constituée de cellulose, de pectines et d’hémicellulose ( voir B. cellulaire). II.1.2.2 Composition de paroiPECTINES :sont
des polymères de plusieurs polysaccharides acides. Les chaînes formées
sont reliées entre elles pour constituer un réseau. les liaisons sont
des laisons faibles qui peuvent facilement se rompre. Il en résulte une certaine élasticité.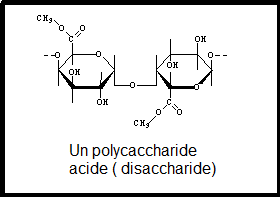 CELLULOSEest un b-1-4 glucane. Elle est insoluble dans la plupart des solvants CELLULOSEest un b-1-4 glucane. Elle est insoluble dans la plupart des solvants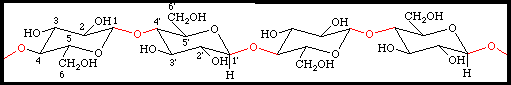 les
chaîne de cellulose forme une structure cristalline appelée
microfibrille qui contient 2000 molécules et qui possède un diamètre de
20 à 30 nm. Il
existe des liaisons intrachaînes qui stabilisent la molécule et des
liaisons interchaînes qui maintiennent les molécules disposées
parallèlement dans les microfibrilles. HEMICELLULOSES constituent la matrice de la paroi.. Ils peuvent être ramifiés et parfois constituer des tissus de réserve ( polymère 1-4 de D xylulose)PROTEINESOn note également la présence de protéines riches en Proline et de glycoprotéines riches en hydroxyproline ( extensines …), celles ci sont nommées protéines pariétalesII.1.2.3 Structure théorique de la cellule végétale jeune La cellule végétale jeune possède donc :Une section polygonale régulièreUn noyau sphérique de type parfait en position centrale dans la cellule.Un cytoplasme ambiant, plus abondant que dans une cellule animale.Une paroi pecto-cellulosique encore
appelée membrane ou paroi primaire constituée essentiellement de
pectine et de cellulose ( voir composition, plus haut) ; mitoyenne, à
contours nets, anguleux ; perméable et souple, ce qui donne à l’ensemble
une section
plus ou moins polygonale. Cette paroi comporte des interruptions au
niveau de la cellulose ( ponctuations : voir schéma). La couche
constituée de pectine est fréquemment appelée lamelle moyenne Remarque : On remarque déjà au niveau des cellules jeunes, la présence de structures appelées plasmodesmes :
ce sont des orifices de communication entre cytoplasmes voisins
contenant des filaments cytoplasmiques, les plasmodesmes laisse passer
en leur centre des canalicules émanant du réticulum endoplasmique,
réalisant ainsi une continuité entre cytoplasmes voisins. [u]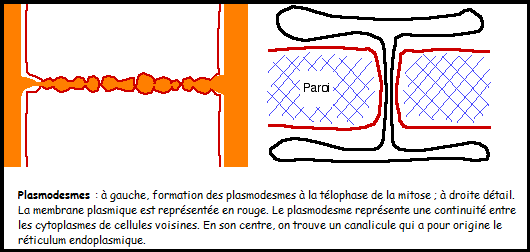 [/u] [u] [/u] [u]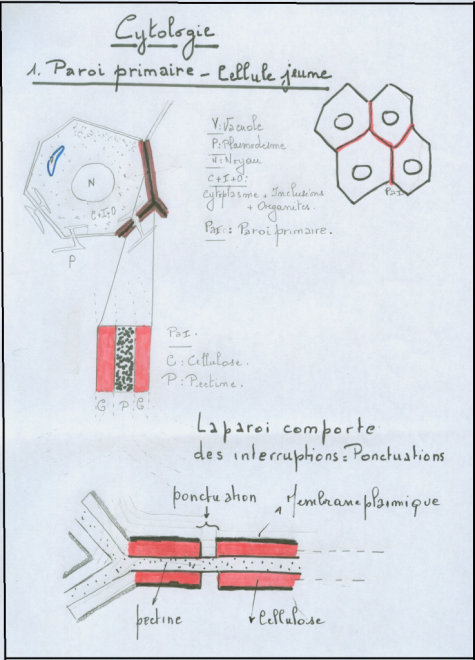 [/u] II.1.3 La cellule adulte C’est une cellule jeune qui a évolué :
- Le noyau est refoulé latéralement, il a perdu sa forme sphérique.
- Le cytoplasme
( partie délimitée par la membrane plasmique et hormis le noyau) ou
compartiment cytoplasmique occupe l’intérieur de la cellule ; on y
distingue une partie transparente hyaline qui a la texture d’un gel ou hyaloplasme apparaissant surtout en périphérie ( ectoplasme des Protozoaires) et une partie très opaque, interne ou endoplasme qui contient les inclusions cytoplasmiques, dont la plus volumineuse est la vacuole.
- Le plasmalemme
ou membrane plasmique délimite la partie externe du hyaloplasme, cette
membrane a d’importantes fonctions physiologiques ( voir B .
cellulaire).
- La paroi primaire ou paroi pectocellulosique est en réalité une enveloppe plus ou moins rigide, servant de contenant au cytoplasme.
La paroi cellulaire apparaîtra avec des contours plus ou moins arrondis aux angles arrondis , on y distingue :Une paroi pecto-cellulosique dite primaire Ou
paroi primitive de la cellule jeune. Elle est en partie rupturée au
niveau de la pectine, selon l’importance de la pression vacuolaire
interne.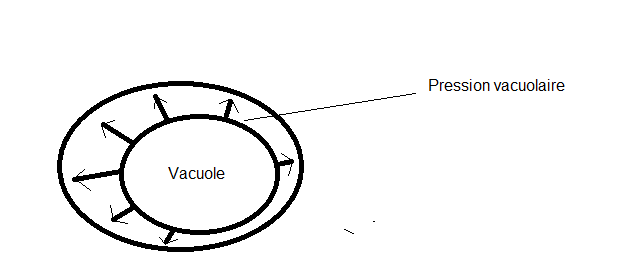 Les Les
couches latérales de cellulose, plus élastiques que la couche de
pectine, on résisté à la rupture grâce à une incorporation de nouvelles
molécules de cellulose ( intussusception).Une paroi pecto-cellulosique dite secondaire Composée
de nouvelles molécules de cellulose apposées par couches successives (
apposition ou placage ) discontinues mais parfois importantes, contre
la membrane primaire. 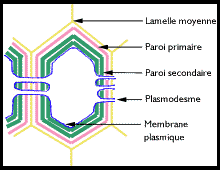 Cette Cette
paroi secondaire pourra dans certains cas se transformer par voie
d’imprégnation ou de recouvrement. Le résultat des transformations
subies par la membrane secondaire donne lieu a des spécialisations
particulières que l’on retrouve dans divers tissus définitifs tels que
le protecteur ( épiderme ) , le soutien ( sclérenchyme), ou le
conducteur ( les vaisseaux de divers types).La transformation de la paroi secondaire par imprégnation se fait au moyen de : - Cutine ( épiderme foliaire ) - Subérine ( liège)- Lignine ( cellules du bois, vaisseaux et fibres). La lignine est un Polyphénol.La transformation de la paroi secondaire par recouvrement se fait au moyen de :- Cire ( Palmiers à cire ) - Matières huileuses ( épiderme d’une pomme ). - Matières minérales ( SIO2 des feuilles de poacées, CaCO3 de certaines algues).Selon le cas , la membrane devient l’élément principal du tissus protecteur ou le tissus squelettique de la plante.En résumé, la paroi est l'enveloppe la plus externe de la cellule végétale. Elle est essentiellement composée de polymères glucidiques,
cellulose et pectine, de protéines pariétales et éventuellement
d'autres composées de nature phénolique (lignine et subérine). La paroi
est composée de trois parties :
- paroi primaire,
de nature pecto-cellulosique, la paroi primaire n'existe seule que dans
les cellules juvéniles. Elle est extensible, ce qui permet la
croissance cellulaire (élongation),
- paroi secondaire,
elle apparaît lors de la différenciation de la cellule. Elle est
constituée de cellulose et d'hémicellulose et est enrichie en composés
phénoliques : lignine (pour renforcer la rigidité), cutine et subérine
(pour l'imperméabiliser).
Cette
différenciation s'observe pour les cellules conductrices de sève du
xylème (le bois) et pour différents tissus de soutien (sclérenchyme) ou
de protection (liège).
- lamelle moyenne,
c'est la partie la plus externe de la paroi et elle est commune à deux
cellules contigües. C'est elle qui se forme la première et elle est
constituée de matières pectiques.
La
paroi assure le maintien et définit la taille et la forme de la cellule
végétale. Elle participe à la régulation des relations avec les autres
cellules et avec l'extérieur, et, de manière passive, au transport, à
l'absorption, et à la sécrétion de multiples substances. Pour permettre
les communications entre cellules, directement de cytoplasme à
cytoplasme, les parois sont finement ponctuées de plasmodesmes.Il faut cependant noter que toute les cellules n’évoluent pas jusqu’au stade recouvrement ou imprégnation …Aux endroits de décollement de la triple paroi pecto-cellulosique apparaissent des méats
ou espaces inter-cellulaires. Lorsque les méats deviennent important,
par convergence, par absorption de cellules voisines ou pour d’autres
raisons, ils sont alors appelés lacunes .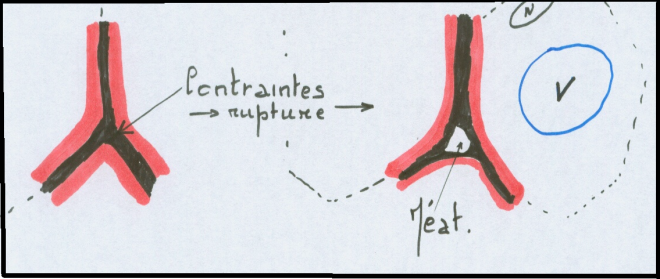 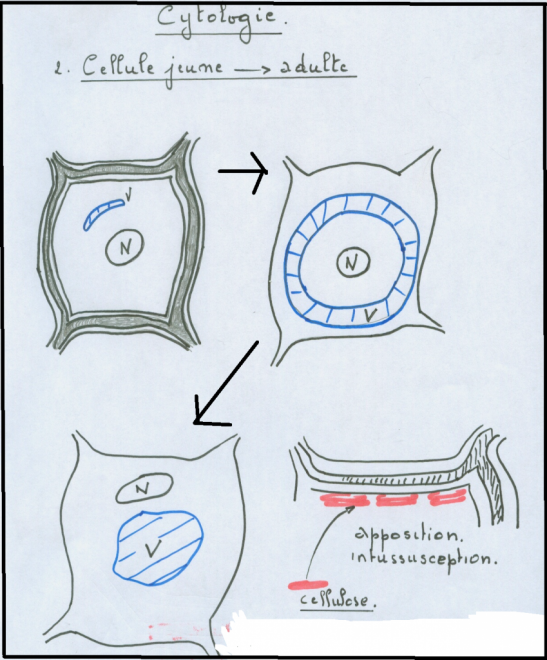 Ces méats contiennent en général des gaz, dont le CO2, l’O2,L’H2O … mais aussi parfois certains liquides de sécrétion ( contenu du Ces méats contiennent en général des gaz, dont le CO2, l’O2,L’H2O … mais aussi parfois certains liquides de sécrétion ( contenu du
zeste d’orange ).Les espaces intercellulaires ont un rôle passif,
néanmoins très important pour certaines fonctions hysiologiques (
photosynthèse, respiration ). La vacuole : La vacuole est une poche contenant en majorité de l’eau qui naît à partir de diverticules de l’appareil de Golgi, par confluences
successives jusqu’au moment ou elle occupe la position centrale dans la
cellule, déplaçant le noyau latéralement. Elle peut toutefois rester
fragmentée.La vacuole est très importante chez les végétaux,elle occupe
80 à 90% du volume cellulaire. Les cellules jeunes possèdent plusieurs
petites vacuoles alors que la cellule adulte se caractérise par une
vacuole unique. La vacuole est limitée par une membrane : le tonoplasme.
Elle contient le suc. La vacuole est très
importante
chez les végétaux, elle occupe 80 à 90% du volume cellulaire. Les
cellules jeunes possèdent plusieurs petites vacuoles alors que la
cellule adulte se caractérise par une vacuole unique. La vacuole est
limitée par une membrane : le tonoplasme. Elle contient le suc
vacuolaire dont la composition varie en fonction de l'état de la plante. En général, son rôle est dédié au stockage :de l'eaude solutés organiques ( sucres en C6 ou C12 ) d'ions minéraux parfois de pigments (anthocyanes) des gaz dissous et parfois des alcaloïdes ( bases organiques azotées comme : nicotine, colchicine,morphine, belladone ) Corps cristallisés comme l’oxalate de Ca ( macle, briques, raphides ) A ce titre, la vacuole joue un rôle majeur dans la régulation des grandes fonctions physiologiques de la cellule végétale (pH, pression osmotique, concentrations ioniques,...). Cette vacuole sert également de réserve d’eau et de réserve alimentaire ( sucres), sa turgescence assure une certaine rigidité aux organes qui ne possèdent pas de tissus de soutien. Les plastes :
sont les inclusions cytoplasmiques les plus caractéristiques de la
cellule végétale. Les cellules végétales contiennent les plastes, des
organites caractéristiques limités par deux membranes, contenant un
stroma, de l'ADN et capables de se diviser à l'intérieur de la cellule.
Ils proviennent de corps incolores très petits, appelés proplastes dans les jeunes cellules indifférenciées, et qui se transforment ensuite en chloroplastes, chromoplastes et leucoplastes,
conformément aux fonctions exprimées par les différents types
cellulaires. Dans une cellule végétale, il est évident qu'il existe une
relation très étroite entre les différents plastes : les chloroplastes
peuvent se transformer en leucoplastes, et inversement, ou en
chromoplastes. 1° Chloroplastes : Les chloroplastes colorent en vert les feuilles et divers organes , tiges jeunes, bractées, stipules, sépales, certaines fleurs, …Structure du chloroplasteUn chloroplaste est composé d'une double membrane isolant un liquide, le stroma, du contenu cellulaire, le hyaloplasme. Un système membranaire, issu de la membrane interne, forme un ensemble de longs sacs aplatis, les thylakoïdes, baignant dans le stroma.L'empilement des thylakoïdes porte le nom de granum.Dans
le stroma on trouve de l'ADN, des ribosomes, des globules lipidiques
(qui permettent la construction des membranes des thylakoïdes) ainsi que
certaines fois des grains d’amidon. La molécule de chlorophylle est
portée par la membrane des thylacoïdes.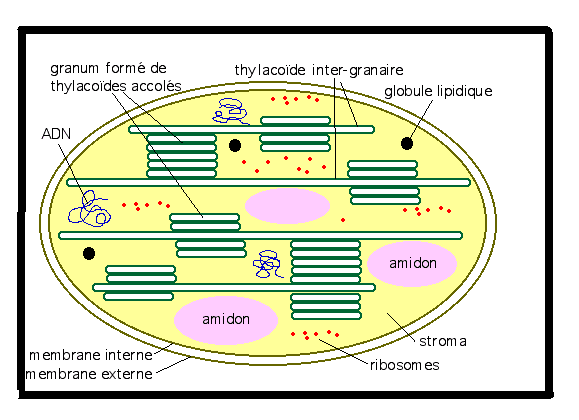 Chez les algues, les chloroplastes sont soit rubanés, en étoile, etc… Chez les végétaux supérieurs, Chez les algues, les chloroplastes sont soit rubanés, en étoile, etc… Chez les végétaux supérieurs,
les chloroplastes sont ovoïdes ou lenticulaires, petits ( 1 à quelques
microns) et nombreux, ils sont d’autant plus petits et nombreux qu’ils
appartiennent à un végétal plus élevé en organisation.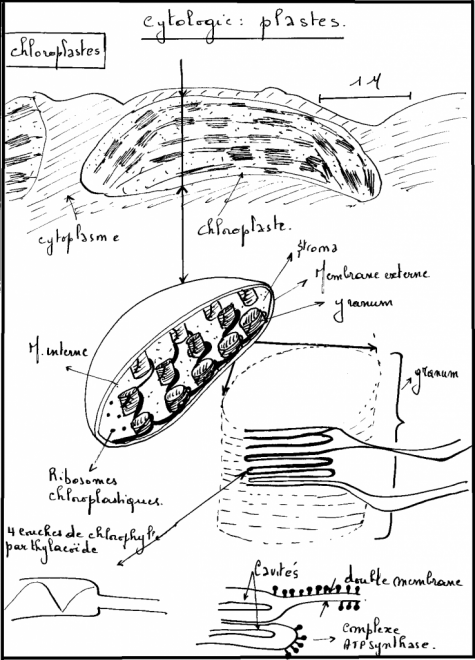 Origine du chloroplaste : la théorie endosymbiotique Dès le début du 20 ème siècle les chercheurs ont pensé que les plastes et les mitochondies pouvaient provenir de bactéries.Celles-ci Origine du chloroplaste : la théorie endosymbiotique Dès le début du 20 ème siècle les chercheurs ont pensé que les plastes et les mitochondies pouvaient provenir de bactéries.Celles-ci
auraient été ingérées par des cellules primitives et vivraient à
l'intérieur d'elles en symbiose. Cette théorie endosymbiotique de
l'origine des plastes et des mitochondries est devenue parfaitement
plausible lorsque l'on a découvert (1950-1960) que ces organites
contenaient de l'ADN et des ribosomes.La
ressemblance entre un chloroplaste de cellule eucaryote actuelle et
d'une bactérie photosynthétique (Cyanobactérie) est confortée par
plusieurs caractères :
l'ADN du chloroplaste est circulaire et non associé à des histones,
comme chez les bactéries,cet ADN code pour une partie des protéines
chloroplastiques (organites semi autonomes),une partie de la synthèse de
protéines chloroplastiques s'effectue dans le chloroplaste, grâce à la
présence de ribosomes qui présentent des analogies avec les ribosomes
bactériens,tout plaste provient d'un plaste préexistant. Lorsque des
cellules ne possèdent pas de plaste (certains cellules blanches de
feuilles panachées), les cellules filles ne possèdent pas de plaste,la
division des chloroplastes suit un rythme indépendant de la division du
noyau, chez lesplantes
supérieures, les deux membranes de l'enveloppe du chloroplaste sont
différentes : la membrane interne ainsi que les membranes des
thylacoïdes présentent des analogies (composition lipidique) avec les
membranes bactériennes.
L'ensemble de ces observations représente aujourd'hui des arguments
forts de la théorie endosymbiotique. Les endosymbioses ont pu se
réaliser à différents moments et de diverses façons, par absorption par
une cellule (Procaryote ou Eucaryote)primitive d'une autre cellule
(Procaryote ou Eucaryote). On parle alors d'endosymbiose primaire ou
secondaire.Endosymbiose primaire (plastes de Rhodophycés et des Chlorophycées)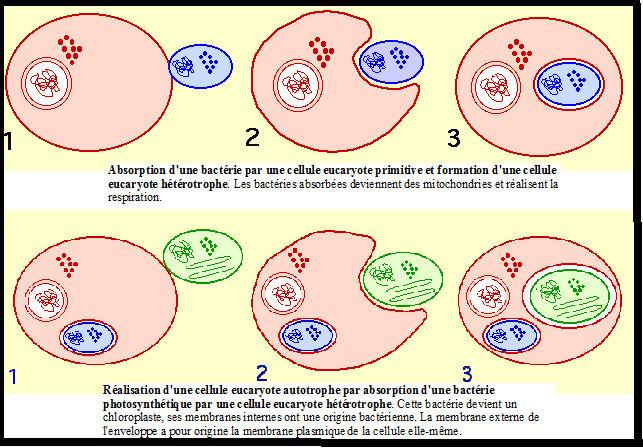 Il Il
est probable que cette endosymbiose ait pu se réaliser de différentes
manières. Chez les algues rouges (Rhodophycées), on constate que les
thylacoïdes possèdent des pigments accessoires, les phycobilines
(phycocyanine et phycoérythrine), ce qui laisse penser que la bactérie
symbiotique devait être une Cyanobactérie qui possédait ces mêmes
pigments. Pour expliquer l'origine des chloroplastes des algues vertes
et des végétaux supérieurs qui contiennent des chlorophylles a et b et
pas de phycobilines, on peut envisager, soit que la Cyanobactérie
symbiote possédait un équipement pigmentaire différent lors de
l'absorption, soit que l'évolution pigmentaire se soit réalisée
ultérieurement.endosymbiose secondaire (plastes des Chromophytes)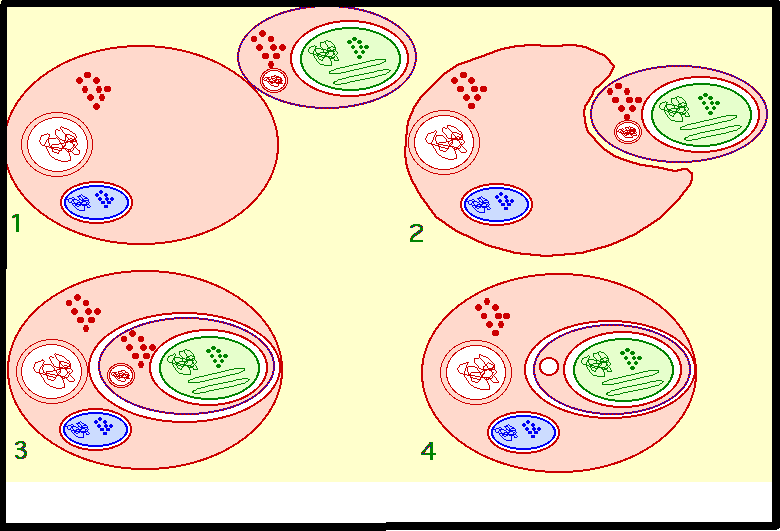 Une Une
cellule eucaryote hétérotrophe absorbe une autre cellule eucaryote
autotrophe contenant un chloroplaste limité par une enveloppe à deux
membranes (endosymbiose primaire). La
membrane plasmique de la cellule symbiote et la membrane de phagocytose
constituent une deuxième enveloppe externe. En général, le noyau et le
cytoplasme de la cellule symbiote dégénèrent, le chloroplaste est alors
entouré de quatre membranes . Chez Cryptomonas (chromophyte,
Crytophycées), on trouve effectivement un reste de noyau (ADN) entre la
deuxième et la troisième membrane (nucléomorphe) ainsi que des restes de
cytoplasme contenant des ribosomes. La fonction chlorophylienne
Nous ne ferons ici qu’effleurer le sujet,celui-ci fera l’objet d’une
étude complète dans la rubrique traitant de physiologie végétale.La fonction chlorophylienne ou assimilation chlorophylienne ou photosynthèse est une caractéristique fondamentale des végétaux certaines bactéries et certains protistes en pratiquent une forme semblable. Définition : Synthèse de glucides ( assimilats ) à partir de l’eau, du CO2atmosphérique
et de l’énergie solaire au niveau d’ un pigment ( chlorophylle ) situé
dans les tissus chlorophylliens ( principalement les feuilles pour les
végétaux supérieurs ). La réaction libére de l’O2 dans l’atmosphère et de l’eau résiduelle. Les organismes photosynthétiques sont pratiquement les seuls capables d’utiliser directement l’énergie solaire.La photosynthèse est donc un acte photochimique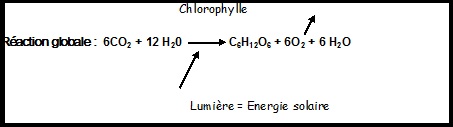 Les deux phases de la photosynthèse La phase lumineuse : permet de capter l’énergie des rayons lumineux et de la transformer en énergie chimique ( NADPH2 Les deux phases de la photosynthèse La phase lumineuse : permet de capter l’énergie des rayons lumineux et de la transformer en énergie chimique ( NADPH2
, ATP…) au niveau des molécules photoréceptrices dont la plus importante est la chlorophylle.
Les molécules photoréceptrices, localisées sur la membrane externe des
thylakoïdes, baignent dans le stroma. L'intérieur des thylakoïdes est
constitué des principaux pigments photosynthétiques (chlorophylle,
caroténoïdes et phycobilines) et des protéines responsables du transport
électronique. La phase obscure: est la phase chimique à proprement parler, elle aboutit à la synthèse de glucides grâce à l’énergie chimique emmagasinée lors de la phase lumineuse ( NADPH2 , ATP…), la photolyse de l’eau et le CO2 ( cycle de Calvin,voir biochimie) .Le
stroma est le lieu où se déroulent les différentes étapes chimiques de
la photosynthèse. Les chloroplastes produisent aussi des protéines grâce
à l'apport de leur propre ADN transmettant l'information génétique
nécessaire. Tout ce processus nécessite des catalyseurs (Mn2+ , Cl- …).Les
glucides produits par la photosynthèse servent soit directement comme
pourvoyeur d’énergie ( mitoses, méiose,absorption racinaire…) soit
constituent des réserves après polymérisation ( amidon…).2° Les amyloplastes (grains d’amidon), les leucoplastes. Amyloplastes
: Les amyloplastes sont des plastes spécialisés dans les fonctions de
réserve. Ils stockent sous forme d’amidon les sucres formés par
photosynthèse jusqu’à ce que la plante en ait besoin. On les trouve dans
les parenchymes spécialisés de certains organes: racines tubérisées,
rhizomes, tubercules, parenchymes du bois (tronc des arbres), quelques
fruits et dans les graines.
Ils sont souvent de forme sphérique, quelquefois lenticulaire, conique
ou polygonale. Leur forme dépend du nombre et du volume des grains
d’amidon accumulés. Les amyloplastes peuvent verdir s'ils sont exposés à
la lumière (sommet de la racine de la carotte, tubercule de pomme de
terre).L’amidon apparaît donc dans un plaste ( stroma) sous l’aspect d’un granule brillant dont le point de départ est appelé hyle
. Il grossit par apposition de couches périphériques nouvelles, d’où
son aspect particulier. Par exemple, le grain d’amidon de la pomme de
terre apparaît avec des stries successives, et l’ensemble rappelle
l’aspect d’une écaille de moule.Bref,
ses aspects sont variés : avec un hyle ( blé, pomme de terre ), deux
hyles ( avoine, riz).Ils se révèlent par la réaction typique qu’ils
présentent avec l’iode ioduré (IKI).Leucoplastes : Ces plastes ont été longtemps regroupés avec les amyloplastes. Ils ont aussi des fonctions de réserve. On les rencontre
dans les cellules sécrétant des essences ou des résines comme les
canaux sécréteurs dans les aiguilles de pin. Certains peuvent se
différencier en chloroplastes ou en chromoplastes s'ils sont exposés à
la lumière, d'autres ne le peuvent pas (leucoplastes des cellules de
garde ou feuilles blanches de certaines variétés de plantes).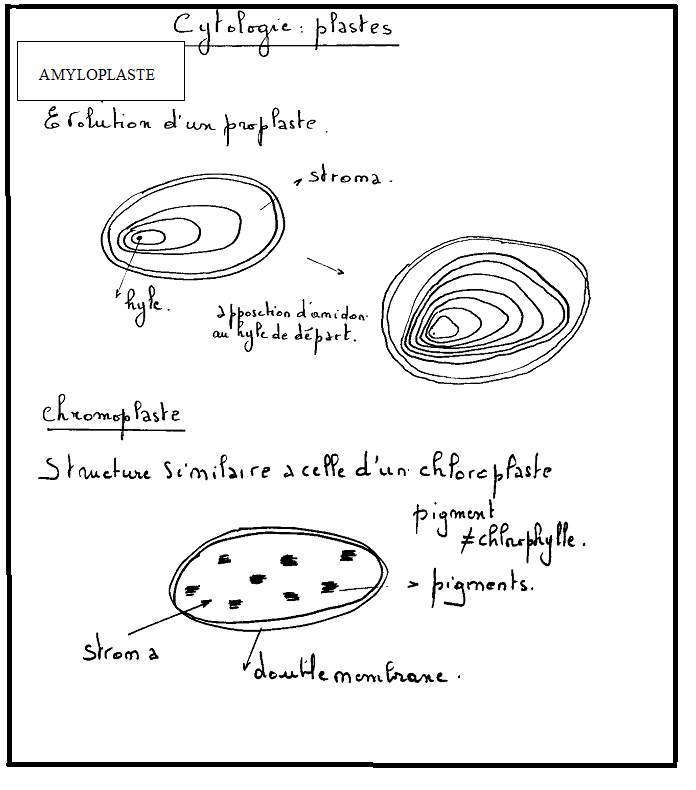 3° Les chromoplastes 3° Les chromoplastes
Les chromoplastes sont des plastes contenant des pigments caroténoïdes,
responsables de la couleur jaune, orangée ou rouge de beaucoup de
fruits (tomate, piment), de pétales de fleurs (jonquille, capucine), ou
même de racines (carotte). Ces plastes accomplissent peu de fonctions
métaboliques. Ils assurent encore, au sein de la cellule végétale, des
fonctions biosynthétiques (lipides, acides aminés, ...), mais, en raison
de la perte des photosystèmes, ils présentent une ultra-structure très
différente de celle des chloroplastes. En général, ils se différencient
tardivement au cours de la maturation des fruits (changement de
couleur), le plus souvent à partir de chloroplastes (tomate, piment,
poivron, etc.). Il ne s’agit toutefois pas d’une règle absolue, car ils
peuvent de former à partir d’amyloplastes (carotte). Leur fonction
semble s’intégrer dans l’évolution des plantes à fleurs où la couleur a
un rôle important à jouer pour assurer la pollinisation donc le succès
de la reproduction.Les éléments les plus caractéristiques sont les structures de stockage des caroténoïdes.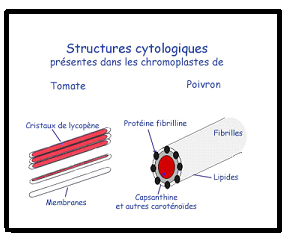 Remarque : Les plastes peuvent évoluer d’une forme à une autre, notamment dans les cellules vieillissantes.Par exemple, les amyloplastes peuvent devenir des chloroplastes ( pomme de terre qui verdit) ou les chloroplastes peuvent se transformer en chromoplastes : chez la tomate qui mûrit ou chez lesfeuilles à l’automne. II.2. Remarques complémentaires - Le noyau de la cellule végétale n’est pas plus développé que celui de la celluleanimale , mais les dimensions générales de la cellule végétale sont en moyenne plus importantes que celles de la cellule animale. - Les cellules animales ne possèdent pas de paroi, il n’y a ni pectine ni cellulose au niveau de la membrane plasmique animale. - La cytocinèse végétale suit un tracé centrifuge, tandis que la cytocinèse animale suit un tracé centripède. - Certains thallophytes possèdent des structures cellulaires particulièresCellulaire : Unicellulaires uninucléés Pluricellulaires : filamenteux, lamellaires ou massifs Symplastique : Cytoplasme indivis multinuclééArticulé : Cytoplasme compartimenté plurinucléé- On distingue trois types de paroi en fonction du degré d’évolution : La paroi primitive : composée de pectine et qui deviendra la lamelle moyenne La paroi primaire : triple couche cellulose – pectine – cellulose mitoyenne avec lamelle moyenne au milieuParoi secondaire : Constituée par placage de cellulose sur la face cytoplasmique de la paroi primaire. Remarque : Les plastes peuvent évoluer d’une forme à une autre, notamment dans les cellules vieillissantes.Par exemple, les amyloplastes peuvent devenir des chloroplastes ( pomme de terre qui verdit) ou les chloroplastes peuvent se transformer en chromoplastes : chez la tomate qui mûrit ou chez lesfeuilles à l’automne. II.2. Remarques complémentaires - Le noyau de la cellule végétale n’est pas plus développé que celui de la celluleanimale , mais les dimensions générales de la cellule végétale sont en moyenne plus importantes que celles de la cellule animale. - Les cellules animales ne possèdent pas de paroi, il n’y a ni pectine ni cellulose au niveau de la membrane plasmique animale. - La cytocinèse végétale suit un tracé centrifuge, tandis que la cytocinèse animale suit un tracé centripède. - Certains thallophytes possèdent des structures cellulaires particulièresCellulaire : Unicellulaires uninucléés Pluricellulaires : filamenteux, lamellaires ou massifs Symplastique : Cytoplasme indivis multinuclééArticulé : Cytoplasme compartimenté plurinucléé- On distingue trois types de paroi en fonction du degré d’évolution : La paroi primitive : composée de pectine et qui deviendra la lamelle moyenne La paroi primaire : triple couche cellulose – pectine – cellulose mitoyenne avec lamelle moyenne au milieuParoi secondaire : Constituée par placage de cellulose sur la face cytoplasmique de la paroi primaire.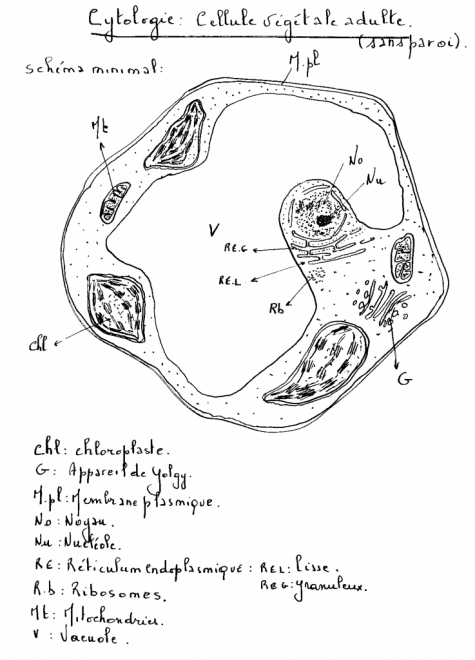 II.3 Rappels concernant les divisions cellulaires1° La mitose II.3 Rappels concernant les divisions cellulaires1° La mitose
est celle qui a pour résultat la production de deux cellules filles à
partir d’une cellule mère : Le caryotype est conservé, les cellules
filles sont identiques à la cellule mère ( même génotype).Exemple, une cellule 2n subit une mitose et donne naissance à deux cellules filles 2nGlobalement, 2n ----------------------> 2 X 2n, on a donc finalement une quantité double de matériel génétique pour les deux cellules.....Comment ?Au cours de l’interphase ( hors mitose) le matériel génétique ( l’ADN) s’est répliqué ( voir la réplication en biologie moléculaire) .Attention, comme nous l’avons dit, par cellule, le matériel génétique est quantitativement ( et qualitativement) conservé 2n ----------> 2nLe processus s’inclut dans le cycle cellulaire et comporte traditionnellement cinq grandes phases : 1) prophase, 2) métaphase,3)anaphase,4)télophase et en 5) la cytocinèse.Nous
ne reviendrons pas sur ce processus, mais notons que contrairement à la
cellule animale, la cellule végétale réalise une cytodiérèse
centrifuge ( formation d’une cloison « sécrétée » par le cytoplasme) et
est dépourvue d’aster et donc de centrioles.L’activité
mitotique est particulièrement abondante dans les tissus jeunes en
croissance ( méristèmes apicaux, des primordiums foliaires, coiffe de la
racine…).2° La méiose
Est la division qui a pour résultat la production de gamètes qui sont
des cellules possédant la moitié de la garniture chromosomique de la
cellule mère.C’est une division typique des tissus germinaux animaux ou
végétaux . Elle permet dans le cadre de la reproduction sexuée de
conserver un génome ayant la même garniture chromosomique au cours des
générations ( sans méiose, le génome doublerait à chaque génération).Exemple : une cellule 2n subit une méiose et donne naissance à quatre cellules filles haploïdesGlobalement, on a : 2n ------------> 4 X n , là encore, la quantité d’ADN a doublé, il y a donc eu réplication préalable.La méiose permet également de conserver la diversité des génomes dans une espèce car : 1) appariemment aléatoire en tétrades au moment de la métaphase 1 2) Phénomène de crossing over (remaniements chromosomiques). La rencontre des gamètes est également réglée par le hasard---------> diversitéDIVERSITE GENETIQUE DE L’ESPECE = SANTE DE l’ESPECE : voir génétique des populations et théories de l’évolution.
III REPRODUCTION ET CYCLES BIOLOGIQUES
On peut définir la reproduction comme suit :
C’est l’ensemble des moyens utilisés pour produire de nouveaux
individus en leur conservant les caractéristiques parentales et celles
de l’espèce.Trois moyens sont utilisés par les plantes : - La multiplication végétative- La reproduction agame - La reproduction sexuée.III.2 REPRODUCTION VEGETATIVEConsiste en la fragmentation d’un individu ou d’une partie de celui-ci.Chaque fragment se développe en un individu nouveau.Les modalités en sont variées :
- Scissiparité ( bactéries) ;
- bourgeonnement ( levures) ; stolon ( = tige rampante , fraisier) ;
- tubercule ( pomme de terre ) ; caieux ( glaïeul, oignon, ail…) ;
- bouture ( peuplier) ; marcotte ( cotoneaster).
Il
est très rare que les plantes annuelles ou bisannuelles présentent une
multiplication végétative naturelle. Cette forme de reproduction
s'ajoute bien à la reproduction sexuée. Quand elle existe elle est
parfois extrêmement efficace pour couvrir rapidement une grande surface
(muguet, fraisier, ronces....). 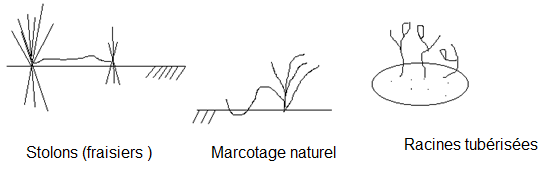 Remarques1) Chez les animaux, seuls certains animaux inférieurs manifestent des potentialités de reproduction végétative ( Spongiaires, Cnidaires, Cténaires, Plathelminthes, Némathelmintes, Annélides,… VOIR Zoologie descriptive).2) Chez les plantes, on rencontre la reproduction végétative à la fois chez les végétaux inférieurs ( Thallophytes, : champignons, lichens…) mais également chez des plantes supérieures. Remarques1) Chez les animaux, seuls certains animaux inférieurs manifestent des potentialités de reproduction végétative ( Spongiaires, Cnidaires, Cténaires, Plathelminthes, Némathelmintes, Annélides,… VOIR Zoologie descriptive).2) Chez les plantes, on rencontre la reproduction végétative à la fois chez les végétaux inférieurs ( Thallophytes, : champignons, lichens…) mais également chez des plantes supérieures.
Beaucoup de plantes supérieures ont des organes de multiplication
végétative spécialisés tubercules,stolons, caieux, rhizomes, drageons…),
elles peuvent également avoir la faculté de produire des racines
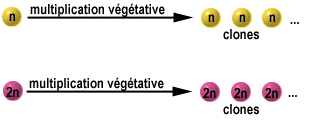
adventives sur des organes normaux ( ex : tiges) d’où plantes nouvelles.3) Notion de lignée végétative ou clone :
C’est l’ensemble des plantes issues d’une même plante mère par voie
végétative . La plante mère de départ s’appelle « tête de clone « ou
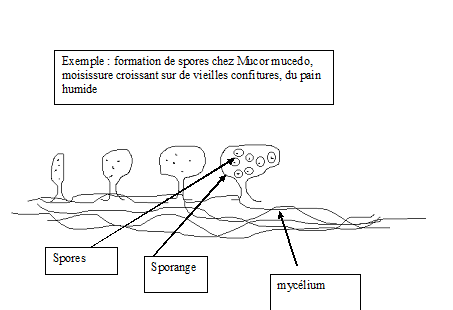
ortet. … applications agronomiquesLA REPRODUCTION AGAME Une
cellule appelée spore est à l’origine d’un nouvel individu, il existe
plusieurs types de spores : endospores, exospores, conidies, zoospores,
etc
III.4 LA REPRODUCTION SEXUEE
Dans la reproduction sexuée, deux cellules sont nécessaires à la
formation d’un nouvel individu. On appelle ces cellules des gamètes,
souvent représentées par la lettre grecque γ , il existe trois types de
reproduction sexuée selon le degré de différenciation entre gamète mâle
et gamète femelle.En isogamie Aucune distinction n’existe entre les gamètes ; cas de certains Thallophytes. Ex : Diatomées.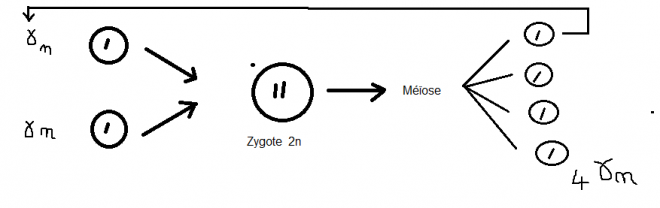 En hétérogamie En hétérogamie
Seule une polarité sexuelle distingue les gamètes ( gamètes + et-, ce
dernier étant immobile par rapport au premier)… certains
thallophytes.Schématiquement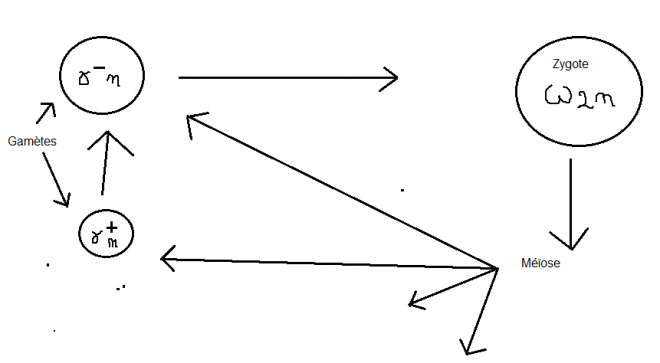 En oogamie,une En oogamie,une
distinction profonde existe entre les gamètes : gamète mâle, mobile,
pourvu d’un flagelle, peu développé, polarisé ; gamète femelle,
immobile, gros, bourré de réserves . Symboles utilisés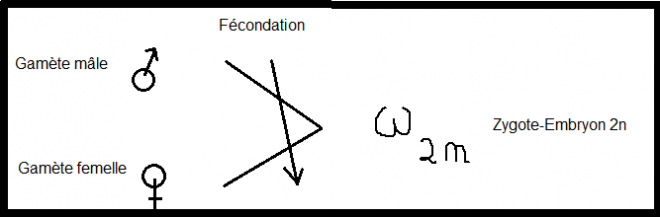 On symbolise la méiose ou cinèse Réductionnelle par : On symbolise la méiose ou cinèse Réductionnelle par : Notons en résumé pour la reproduction sexuée : La reproduction sexuée met en oeuvre deux processus fondamentaux : la méiose et la fécondation. Lors de la fécondation, l'union de Notons en résumé pour la reproduction sexuée : La reproduction sexuée met en oeuvre deux processus fondamentaux : la méiose et la fécondation. Lors de la fécondation, l'union de
deux gamètes haploïdes donne naissance à un zygote diploïde (n->2n)
dont le patrimoine génétique est le résultat de la recombinaison au
hasard des deux génomes parentaux qu'apportent chacun des gamètes. Cas des trois embranchements supérieurs , Bryophytes, Ptéridophytes, Spermatophytes.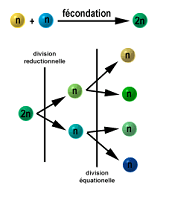 Inversement,au moment de la méiose, Inversement,au moment de la méiose,
le stock chromosomique diploïde est partagé en deux parties
numériquement égales (2n->n), mais qualitativement différentes. Les
individus qui proviennent d'une reproduction sexuée sont donc le
résultat d'un double brassage génétique. Ce sont des individus originaux, génétiquement uniques,
dont certains seront mieux adaptés que d'autres à un nouvel
environnement. La reproduction sexuée est donc le véritable moteur de
l'évolution des êtres vivants et elle permet la création de nouvelles
espèces.III.5 L’ALTERNANCE DES PHASES : LES GRANDS CYCLES BIOLOGIQUESL'alternance
méiose-fécondation introduit un cycle dans le développement d'un
organisme caractérisé par une alternance de phases chromosomiques,
l'haplophase et la diplophase.
Chez les animaux, hormis quelques protozoaires, seuls les gamètes sont
haploïdes,toutes les autres cellules : le zygote, l'embryon et l'adulte
sont diploïdes. Les animaux ne présentent donc au cours de leur cycle
biologique qu'une seule génération diploïde (une seule forme biologique)
et la phase chromosomique haploïde est réduite aux gamètes.Chez
les végétaux, à l'alternance de phases chromosomiques se superpose une
alternance de formes biologiques ou de générations. Toute espèce vit
typiquement et successivement sous deux formes biologiques distinctes,
l'une haploïde correspondant à l'haplophase, l'autre diploïde
correspondant à la diplophase.Le
moment ou se produisent les cinèses réductionnelles n’est donc pas le
même chez tous les végétaux, de sorte que l’on peut distinguer :cycle monogénétique diplophasique,
on a un individu diploïde qui donne naissance à des gamètes haploïdes
par méiose. L'union de ces gamètes donne un zygote diploïde qui par
mitoses successives donne naissance à un nouvel individu diploïde. La
phase chromosomique haploïde est réduite aux gamètes et on a une seule
génération qui est un gamétophyte diploïde,
si on se réfère uniquement à l'éthymologie du mot gamétophyte (= plante
à gamètes). Par contre, si on prend en considération l'état de ploïdie
de la cellule, cette génération unique serait un sporophyte diploïde et
la gamétophyte haploïde aurait disparu, les cellules méïotiques
(=spores) se"transformant" directement en gamètes. Un tel cycle est
typique du règne animal, sauf de quelques protozoaires.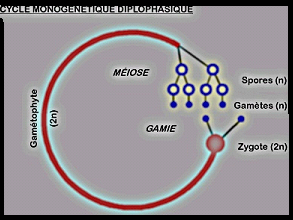 Exemple chez les plantes : Certains Thallophytes ( Diatomées)Dans le cas du cycle monogénétique haplophasique, Exemple chez les plantes : Certains Thallophytes ( Diatomées)Dans le cas du cycle monogénétique haplophasique,
on a un gamétophyte haploïde qui donne des gamètes. La fusion de deux
gamètes mâle et femelle conduit à la formation d'un zygote diploïde qui
va donner par méiose des spores haploïdes.Dans ce cas, la phase
chromosomique diploïde est réduite au zygote. La méiose a lieu
directement dans le zygote.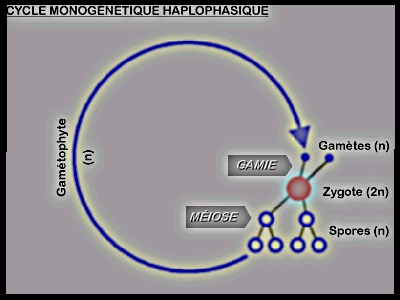 Le cycle digénétique Haplodiplophasique Typiquement Le cycle digénétique Haplodiplophasique Typiquement
aux deux périodes de ce cycle ( haplophase et diplophase) correspondent
deux sortes d'organismes, deux générations. La première est constituée
de cellules haploïdes (n chromosomes). Au sein de ses organes
reproducteurs, elle élabore par simples mitoses des cellules
reproductrices à n chromosomes : les gamètes.Cette génération est donc le gamétophyte (végétal qui génère les gamètes).La
fusion d'un gamète mâle et d'un gamète femelle (=fécondation ou gamie)
donne naissance à un oeuf (=zygote) diploïde dans les divisions
successives par mitose sont à l'origine d'un organisme à cellules
diploïdes qui représentent la seconde génération. Celle ci forme par
méïsose (réduction chromatique) dans ses
organes reproducteurs des cellules haploïdes appelés spores méiotiques ou tétraspores ou méiospores. Cette génération est donc le sporophyte qu'il est préférable de nommer tétrasporophyte ou méiosporophyte. Chacune des spores, par mitoses successives, se développera en un nouveau gamétophyte haploïde.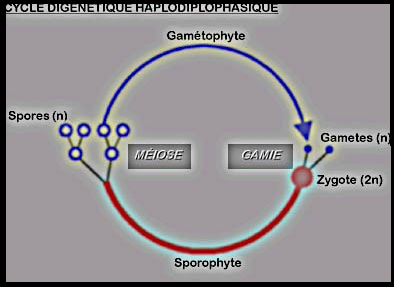 Les Les
cycles digénétiques sont les plus nombreux et les plus diversifiés
puisqu'on les rencontre chez de très nombreux thallophytes et qu'ils
sont les seuls actuellement connus chez les Cormophytes. Ils sont soit
isomorphes chez certains Thallophytes, soit le plus souvent
hétéromorphes chez d'autres Thallophytes et chez tous les Cormophytes
avec prédominance soit de la phase haploïde chez les Bryophytes (cycle
digénétique haplodiplophasique) soit de la phase diploïde chez les Trachéophytes (cycle digénétique diplohaplophasique). Le végétal qui présente un tel cycle est un haplodiplonte ou un diplohaplonte. | |
|

