biologista

Sexe : 
 Messages : 180 Messages : 180
 Points : 317 Points : 317
 Date de naissance : 07/05/1992 Date de naissance : 07/05/1992
 Age : 32 Age : 32
 Emploi : Chez mon papa Emploi : Chez mon papa
 | Sujet: HISTOLOGIE VEGETALE 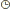 Jeu 6 Déc 2012 - 0:19 Jeu 6 Déc 2012 - 0:19 | |
| source: biodis HISTOLOGIE VEGETALE L’histologie est l’étude des tissus, formés d’un assemblage de cellules, dans notre cas : les tissus végétaux.IV.1 DIFFERENTS TYPES DE TISSUS VEGETAUX1) Le tissus méristématique ou méristème 2) Le tissus de soutien ou squelettique : le colenchyme, le sclérenchyme,les fibres. 3) Le tissus de conduction ou conducteur avec deux types puisqu’il existe deux sèves. 4) Le tissus de protection ou protecteur : l’épiderme, les stomates, les poils épidermiques, le tissus subérisé. 5) Le tissus de sécrétion ou sécréteur : cellules sécrétrices, lacunes, canaux sécréteurs, tubes à latex . 6) Le tissus de remplissage ou parenchyme ou encore tissus parenchymateux.Tableau : les tissus végétaux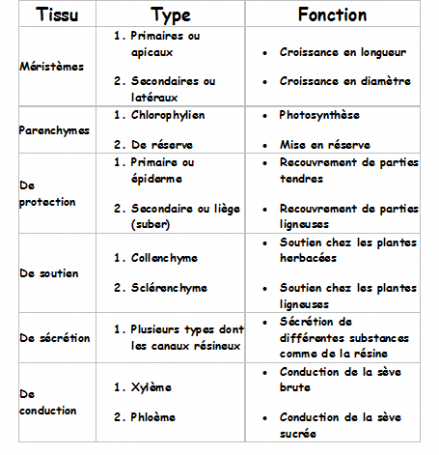 IV.2 CARACTERISTIQUES DES TISSUS VEGETAUX IV.2 CARACTERISTIQUES DES TISSUS VEGETAUX
Il faut noter que chez les plantes, les tissus résultent de la
différenciation des cellules qui proviennent des méristèmes. Ce
processus de formation de cellules spécialisées aptes à remplir une
fonction précise à partir des méristèmes s’appelle différenciation
cellulaire (ce terme est synonyme de spécialisation ou de vieillissement
cellulaire). Les cellules issues de la prolifération des cellules
méristématiques se transforment progressivement en tissus adultes. Au
cours de cette transformation, il en résulte la réalisation de tissus
différents, les cellules acquièrent une forme, une structure et une
physiologie caractéristiques de chaque sorte de tissu. Le phénomène de différenciation cellulaire est caractérisé par :- Un grandissement cellulaire. C’est cette élongation qui est responsable de la croissance en longueur de la tige et de la racine.- Un développement important de l’appareil vacuolaire.- Transformation des plaste primitifs en plastes spécialisés (chloroplastes, amyloplastes, etc.).-
Modifications importantes de l’épaisseur de la membrane cellulosiques
et de ces caractéristiques chimiques (souvent, cela entraînera la mort
de la cellule).- Perte de la capacité de division cellulaire (les cellules ne peuvent plus se diviser comme dans les méristèmes).- Adaptations à des fonctions précises: photosynthèse, mise en réserve, protection, soutien conduction, etc.IV.3 TISSUS MERISTEMATIQUELes méristèmes sont des tissus formés par un ensemble de cellules jeunes et indifférenciées, qui se multiplient activement
et rapidement durant la saison végétative. Ils sont à l’origine de tous
les autres tissus de la plante.Ils sont responsables de la croissance indéfinie qui caractérise des végétaux comme les arbres. Chez beaucoup de plantes, il existe une double croissance : une croissance en longueur ou primaire qui permet l’allongement des tiges et des racines et leur ramification, et une croissance en épaisseur ou secondaire qui assure une augmentation en diamètre des tiges et racines. Par conséquent, on distingue deux sortes de méristèmes,
différents l’un de l’autre par leur localisation dans la plante,leurs
caractères cytologiques et leur rôle dans la construction des organes : les méristèmes apicaux ou primaires et les méristèmes latéraux ou secondaires appelés également cambiums.
Cependant,
les plantes ont des propriétés que les animaux n’ont pas généralement.
En effet, certaines cellules végétales spécialisées peuvent se dédifférencier,
c’est-à-dire qu’elles peuvent, dans certaines circonstances, retrouver
leur état méristématique et leur capacité de se diviser. Nous appelons
ce phénomène dédifférenciation cellulaire. Cette capacité qu’ont les
cellules végétales à retrouver leur état méristématique, nous
l’utilisons fréquemment dans la multiplication végétative des végétaux
comme dans le bouturage ou la culture ' in vitro'.
On sait que le bouturage consiste à prélever sur une plante un fragment
de tige et à le planter dans un sol humide; des racines apparaissent
alors à la base du rameau, reconstituant ainsi un organisme végétal
complet. Lorsqu’on étudie comment se forment ces racines on voit
qu’elles résultent d’une dédifférenciation de certaines cellulesspécialisées
de l’écorce de la tige. Ces cellules retrouvent alors leur capacité de
se diviser qu’elles avaient perdue en se spécialisant. Ce ne sont pas
toutes les cellules qui peuvent se dédifférencier: les parenchymes,
l’épiderme, le collenchyme, le phelloderme et la cellule compagne le
peuvent,les
autres n’ont pas cette capacité, soit que les cellules sont mortes,
aient perdu leur noyau ou que le dépôt de substance sur leur membrane
soit tropimportant.Méristèmes primaires ou apicaux Les méristèmes primaires ou apicaux, situés à l’extrémité (apex) des racines et dans les bourgeons des pousses axillaires et de la tige, fournissent les cellules nécessaires à la croissance en longueur et à la ramification. Les tissus d'un embryon dans une graine sont constitués par du méristème qui, donnera lorsque la graine germe, tous les tissus de la nouvelle plante.
Cette phase de multiplication cellulaire est la première étape dans
l’ensemble des processus qui assurent l’allongement de la racine et de
la tige (la deuxième étape étant l’allongement des cellules
elles-mêmes). Ce type ’élongation porte le nom de croissance primaire
et l’ensemble des cellules issues de ce processus, tissus primaires.
Les plantes herbacées ne connaissent que la croissance primaire tandis
que les plantes fortement lignifiées ont en plus une croissance
secondaire issue de méristèmes secondaires.Les
cellules des méristèmes primaires sont petites, isodiamétriques,
c’est-à-dire ne présentant aucun allongement dans une direction
privilégiée, et parfaitement jointives (pas d’espaces entre les
cellules). La membrane cellulosique est très mince, les vacuoles
abondantes et petites et le noyau volumineux. Il n’y a pas d’axe précis
de division cellulaire. Aujourd’hui, nous mettons à profit cette
capacité illimitée de division cellulaire dans la culture “in vitro” de
méristèmes. Méristèmes secondaires ou latéraux (cambiums) Les méristèmes secondaires sont localisés dans les parties âgées des tiges et des racines dont ils assurent la croissance en diamètre. Ils produiront les tissus secondaires
de la plante. On retrouve ces méristèmes chez les Gymnospermes et les
Angiospermes dicotylédones (ils sont absents chez les angiospermes
monocotylédones, ce qui implique que nous retrouvons rarement des arbres
et des arbustes dans ce groupe). Les cellules des méristèmes secondaires sont rectangulaires disposées en une seule couche très régulière
(on les appelle souvent assises), elles sont allongées suivant le grand
axe de l’organe où elles sont situées et souvent aplaties dans le sens
radial, l’appareil vacuolaire est plus développé que chez les méristèmes
primaires ce qui rejette le noyau à la périphérie. Le noyau est relativement petit. Les cellules se divisent selon un axe tangentiel. 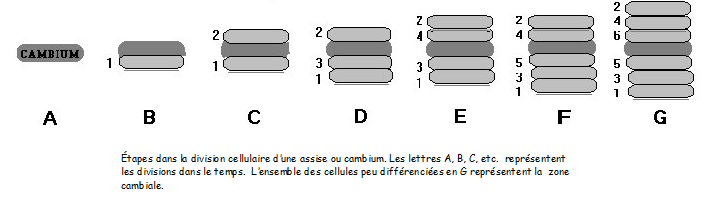 IV. 4 LE PARENCHYME IV. 4 LE PARENCHYME C’est le tissus le moins évolué après le tissus méristématique, puisqu’il réalise avant tout un rôle de remplissage.Les Parenchymes sont des tissus constitués de cellules vivantes, à paroi pecto-cellulosique mince, perforées de ponctuations ou plasmodesmes, C’est le tissus le moins évolué après le tissus méristématique, puisqu’il réalise avant tout un rôle de remplissage.Les Parenchymes sont des tissus constitués de cellules vivantes, à paroi pecto-cellulosique mince, perforées de ponctuations ou plasmodesmes,
qui permettent des communications intercellulaires et une circulation
des substances à l'intérieur des cellules (circulation symplastique).Les cellules peuvent être jointives, collées par leur lamelle moyenne; mais elles présentent
souvent des décollements liés à la gélification de la lamelle moyenne
dans les angles (parenchyme à méats), pouvant aller jusqu'à la formation
d'espaces importants (parenchyme lacuneux). Ces espaces, ainsi que les
propriétés de la paroi pecto-cellulosique permettent une circulation des substances à l'extérieur des cellules et dans la paroi (circulation apoplastique de l'eau, des substances dissoutes et de gaz. Composition : Cellules vivantes noyau + cytoplasme ; vacuole plus ou moins développée ; membrane pecto-cellulosique à peine épaissie ; plastes présents ou non. Les Les
divers types de parenchymes se caractérisent selon les inclusions
qu’ils contiennent, et qui leur attribuent une seconde fonction, souvent
plus importante que le remplissage. - Le parenchyme vert ou chlorophyllien
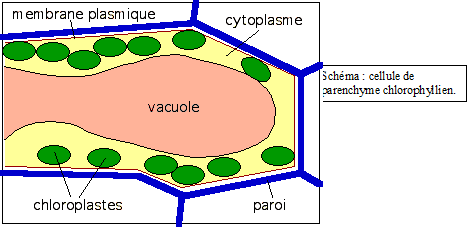 Ce type de parenchyme est pourvu de chloroplastes, il est méatique et vacuolisé.On le rencontre dans tous les organes verts. - Parenchyme blanc ou à leucoplastes. Légèrement méatique, il se situe dans les organes et tissus de réserve Ce type de parenchyme est pourvu de chloroplastes, il est méatique et vacuolisé.On le rencontre dans tous les organes verts. - Parenchyme blanc ou à leucoplastes. Légèrement méatique, il se situe dans les organes et tissus de réserve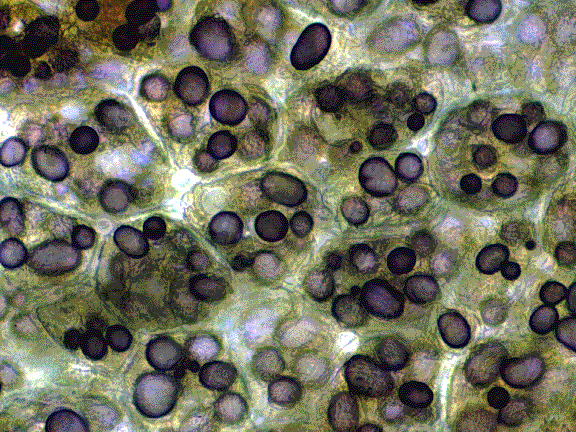 - Parenchyme vacuolaire Avec vacuoles très développées . Par exemple, le parenchyme sucré ( canne à sucre, betterave, fruits charnus ). - Parenchyme améatique et incolore C’est le véritable parenchyme de remplissage, on le rencontre localement dans certains tissus.- Autres parenchymes Aquifère ( plantes succulentes) ; aérifère ( plantes aquatiques ), etc… Voir applications microscopiques IV.5 LE SQUELETTE DE LA PLANTELe - Parenchyme vacuolaire Avec vacuoles très développées . Par exemple, le parenchyme sucré ( canne à sucre, betterave, fruits charnus ). - Parenchyme améatique et incolore C’est le véritable parenchyme de remplissage, on le rencontre localement dans certains tissus.- Autres parenchymes Aquifère ( plantes succulentes) ; aérifère ( plantes aquatiques ), etc… Voir applications microscopiques IV.5 LE SQUELETTE DE LA PLANTELe
tissus parenchymateux en est déjà un puisqu’il réalise la turgescence
cellulaire , mais il n’est pas suffisant pour assurer cette fonction chez des plantes de grande dimension.La plante assure son soutien au niveau de sa membrane secondaire qu’elle épaissit ( cellulose), c’est le collenchyme qu’elle durcit (lignine ) : c’est le sclérenchyme ; ou qu’elle encastre : par les fibres.IV.5.1 Le collenchyme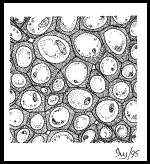 Les Les
épaississements de cellulose de la paroi des cellules du collenchyme
apparaissent sous différentes formes : en pilastres, en barres
biconcaves en anneau. Les cellules du collenchyme sont vivantes
et améatiques, elles fournissent un soutien souple car la paroi
secondaire reste cellulosique. On les rencontre, par exemple dans les
organes mous ( feuilles au voisinage des nervures, tiges herbacées, etc
…), ou dans des organes en croissance.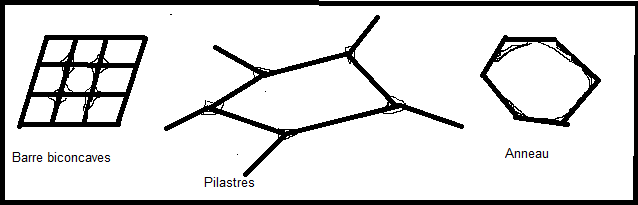 IV.5.2 Le sclérenchyme IV.5.2 Le sclérenchyme 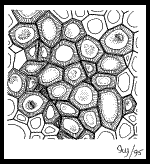 Le Le
sclérenchyme est un parenchyme durci, la paroi secondaire est plus ou
moins développée ( cellulose ) et cette cellulose est imprégnée par de
la lignine . Les cellules scléreuses sont mortes, celles ci sont polygonales ou arrondies. Exemples :
coquilles ( noix, noisettes, noix de coco…), cellules « pierreuses »
(dans certains fruits charnus), Noyau de fruit ( cerise).Des canalicules existent entre les lumens des cellules voisinesNB : existence d’un cas intermédiaire : Le parenchyme scléreux.IV.5.3 Les fibres Constituées de cellules mortes qui réunissent les étapes précédentes de formation du squelette cellulaireEpaississement ( cellulose), durcissement ( lignine), plus l’encastrement : cellules fusiformes qui s’entrelassent Ces Ces
fibres sclérites sont caractérisées par leur longueur en général
importante ( fibres textiles), ainsi que leur degré de lignification (
souplesse ou rigidité).IV.6 LE CONDUCTEUR Il existe deux sortes de sève : 1) La sève brute , minérale, elle est dite « montante » et circule par les vaisseaux. 2) La sève élaborée, organique, elle est dite « descendante » et circule par les tubes criblésIV.6.1 . Les vaisseaux Ce sont de longues cellules mortes, superposées bout à bout, de section circulaire ou polygonale.
Il existe deux grands types de vaisseaux selon que la cloison latérale
est disparue ( trachées ou vaisseaux ouverts ) ou que cette cloison
latérale est maintenue ( trachéides ou vaisseaux fermés).Dans le premier cas, la circulation de la sève est directe et se rencontre chez les Angiospermes ; 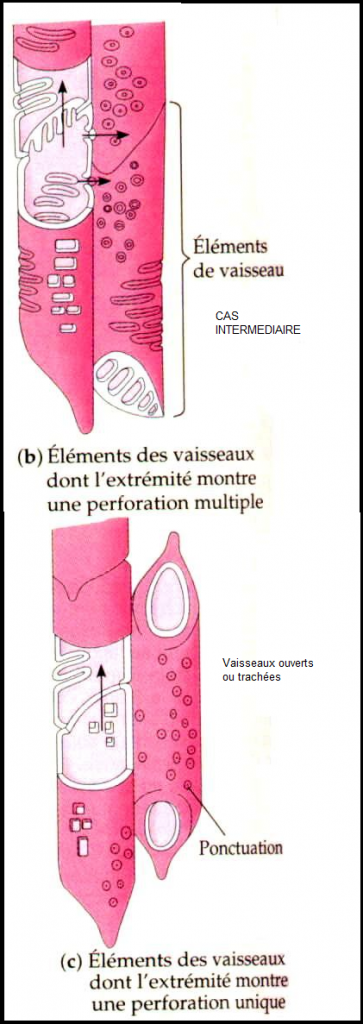 dans le second cas , la circulation est indirecte, se réalise en chicanes , et existe chez les gymnospermes. dans le second cas , la circulation est indirecte, se réalise en chicanes , et existe chez les gymnospermes.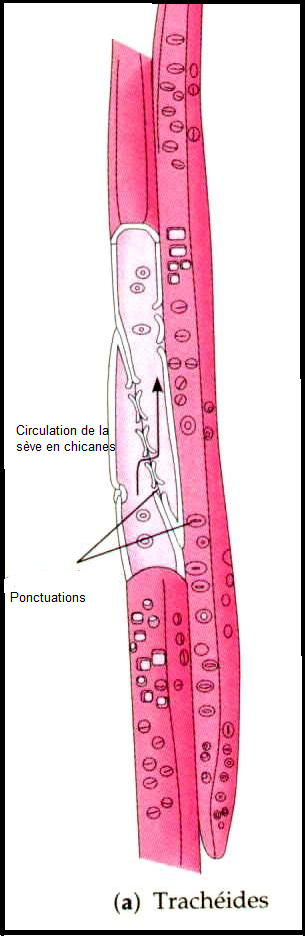 Il faut noter que les trachées comme les trachéides possèdent des ponctuations ( vaisseaux ponctués)sur leur paroi latérale, permettant la circulation latérale de la sève minérale.Remarques 1) chez les fougères, les vaisseaux ont la caractéristique d’être de type scalariforme : les motifs de renforcement ont la forme d’une échelle. 2) Il y a une grande similitude entre les trachéides et les fibres ( sclérites), surtout au niveau de la paroi.Structure des vaisseaux : seule persiste la membrane, mais celle-ci est renforcée , épaissie de diverses manières ( motifs de lignine + cellulose) . D’où existence de divers types de vaisseaux : annelés, réticulés, triés, spiralés, ponctués ( parois recouvertes de lignine sauf aux endroits de ponctuation),Schéma : vaisseau annelé Il faut noter que les trachées comme les trachéides possèdent des ponctuations ( vaisseaux ponctués)sur leur paroi latérale, permettant la circulation latérale de la sève minérale.Remarques 1) chez les fougères, les vaisseaux ont la caractéristique d’être de type scalariforme : les motifs de renforcement ont la forme d’une échelle. 2) Il y a une grande similitude entre les trachéides et les fibres ( sclérites), surtout au niveau de la paroi.Structure des vaisseaux : seule persiste la membrane, mais celle-ci est renforcée , épaissie de diverses manières ( motifs de lignine + cellulose) . D’où existence de divers types de vaisseaux : annelés, réticulés, triés, spiralés, ponctués ( parois recouvertes de lignine sauf aux endroits de ponctuation),Schéma : vaisseau annelé Schéma vaisseau spiralé Schéma vaisseau spiralé 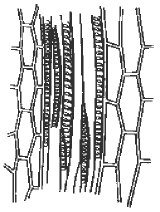 Schéma (2) : comparaison des principaux renforts de vaisseaux chez les angiospermes Schéma (2) : comparaison des principaux renforts de vaisseaux chez les angiospermes 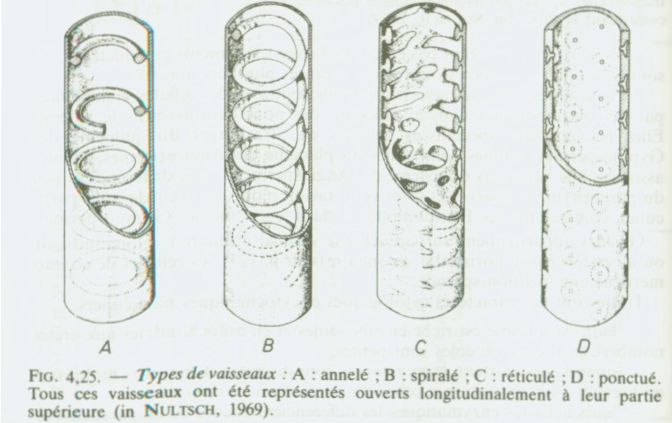 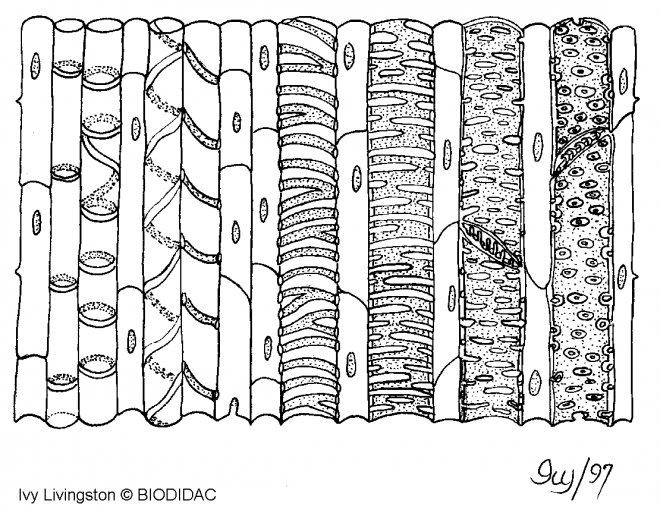 IV.6.2 LES TUBES CRIBLES IV.6.2 LES TUBES CRIBLES
Les tubes criblés assurent la circulation de la sève descendante,tant
entendu que cette expression n’a de sens que si l’on considère le
mouvement de la sève des feuilles vers les racines, mais la sève circule
également vers les bourgeons, ou elle remonte.Les
tubes criblés sont également composé de cellules superposées bout à
bout, mais les membranes transversales persistent et sont perforées.
D’où le nom de cribles donné à ces membranes et le nom de tubes criblés
aux cellules. Il n’existe qu’un seul type de tube criblé, mais les
lumens ont des dimensions variables.
Les tubes criblés sont des cellules vivantes, mais profondément
altérées ; elles sont dépourvues de noyau, le cytoplasme est pariétal et
surtout concentré à la face inférieure, avec une vacuole centrale peu
visible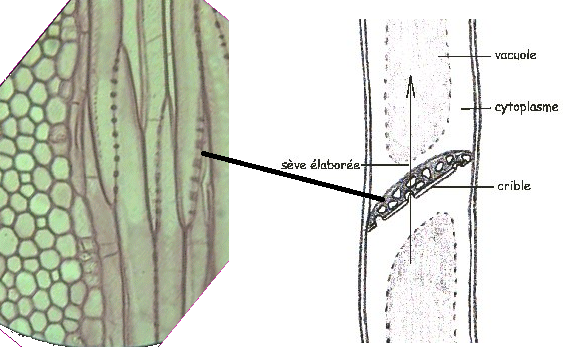 Le cytoplasme comporte également une inclusion volumineuse appelée corps protéino-lipidique dont le rôle est mal connu. Une membrane secondaire tapisse la paroi primaire pecto cellulosique et est plus ou moins lignifiée. Chez les angiospermes, Le cytoplasme comporte également une inclusion volumineuse appelée corps protéino-lipidique dont le rôle est mal connu. Une membrane secondaire tapisse la paroi primaire pecto cellulosique et est plus ou moins lignifiée. Chez les angiospermes,
Chez les angiospermes, les tubes criblés sont flanqué d’une cellule
latérale appelée cellule compagne, pourvue d’un noyau et d’un cytoplasme
, avec perforations latérales au niveau des parois communes avec le
tube criblé. Les cellules compagne jouent des rôles importants,
chargement de glucides dans le tube criblé, réparation des
détériorations au niveau des TC (thylles) etc… voir physiologie
végétale.Schéma : cellule criblée – cellule compagne. 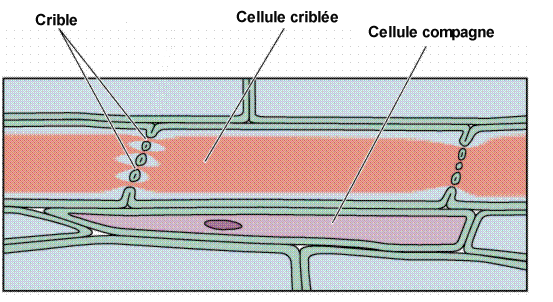 Les Les
tubes criblés sont en contact avec le parenchyme foliaire par des
cellules de transfert assurant la liaison conducteur – parenchyme
chlorophyllien. IV.7 LE PROTECTEUR La plante doit se protéger contre l’atmosphère plus ou moins sèche dans laquelle elle vit, et qu’elle exploite ( CO2, O2
) . Cela justifie l’existence d’un manteau protecteur ( épiderme )
percé d’ouvertures réglables ( stomates) afin de permettre les échanges
gazeux dans des conditions déterminées, compatibles avec les conditions
atmosphériques externes.IV.7.1 Prenons l’exemple d’un épiderme foliaire
Il se compose d’une assise de cellules régulières vivantes ( N + Cy )
mais dépourvue de vacuoles de plastes et de méats ( étanchéité). Son
rôle est exclusivement protecteur et il est assuré par la seule paroi externe : cellulose et substances d’imprégnation ou de recouvrement , de sorte qu’elle reste résistante , souple et imperméable .Ces substances peuvent être constituées de :- Cutine, l’ensemble prend le nom de cuticule ou couche cuticulaire. La cuticule, édifice supramoléculaire impliqué dans la protection des organes végétaux est constituée par une matrice polymérique,
la cutine, polyester d'acides gras hydroxylés et de gycérol dans
laquelle sont inclus des cires et d'autres composés hydrophobes. La
cuticule contrôle les mouvements d'eau entre les organes végétaux .- Cires, ce sont des esters d'acides gras et d'alcools gras, présentes en quantités particulièrement importantes au niveau des tiges de palmier brésilien, de la peau de pomme, prunes…- Sels minéraux : CaCO3, ( algues), SiO2 (graminées,cypéracées ).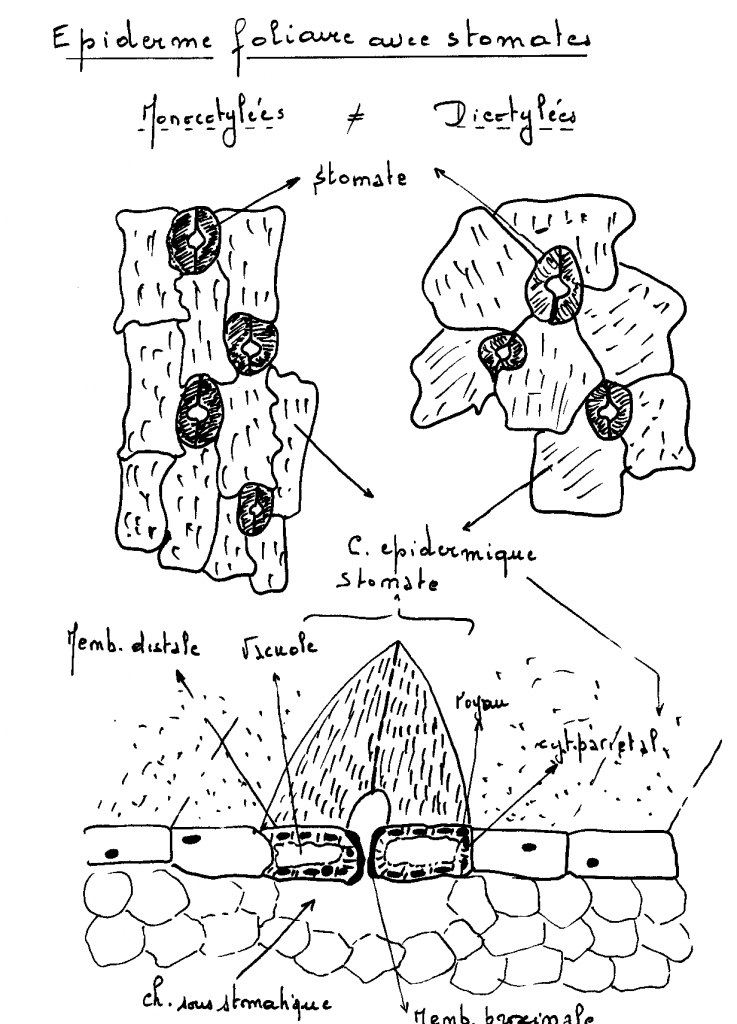 Les épidermes des plantes monocotylées sont constitués de cellules plus longues que larges , quoique de forme variable ; au contraire, Les épidermes des plantes monocotylées sont constitués de cellules plus longues que larges , quoique de forme variable ; au contraire,
les cellules des épidermes de dicotylées sont des cellules
isodiamétrales, avec des contours non réguliers, parfois de forme
analogue aux pièces d’un jeu de puzzle. Cette différence est importante,
mais dans tous les cas, les cellules de l’épiderme sont imperméables et
améatiques.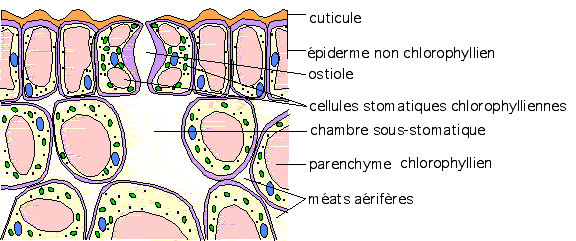 IV.7.2 Les stomates IV.7.2 Les stomates cellules de garde » accolées et ménageant entre elles une ouverture entre elles . Cette ouverture est appelée ostiole. Dans cellules de garde » accolées et ménageant entre elles une ouverture entre elles . Cette ouverture est appelée ostiole. Dans
tous les cas, il se situe dans l’épiderme, où il est encastré ( cas des
mono et dicotylées) ; le stomate peut cependant être relégué au niveau de l’assise sous jacente à l’épiderme ( cas des gymnospermes : coniférales). Voir en anatomie. Les cellules de garde du stomate sont constituées de :
Noyau+cytoplasme+chloroplastes+leucoplastes + vacuole +membrane distale
et membrane proximale ; chambre sous stomatique sous jacente Rem : la membrane proximale est plus épaissie que la membrane distale.Les
stomates sont donc constitués de cellules très complètes, autonomes (
chloroplastes, leucoplastes ), assurant par leur compression ou leur
expansion ( turgescence et plasmolyse des vacuoles) les connexions
gazeuses entre le milieu externe ( atmosphère) et le milieu interne de la feuille ( méats).
Les mécanismes déterminant les mouvement des cellules de garde sont
complexes et font encore l’objet de spéculations ( effet de la lumière,
transports actifs, entrée de K+…). Ces mécanismes sont à étudier en
physiologie végétale.IV.7.3 REMARQUES 1°
La chambre sous stomatique permet de réaliser les échanges gazeux en
deux temps, ce qui les ralentit ; elle est aussi une réserve gazeuse ;
enfin, elle permet de limiter les compressions et expansions de la
feuille.2°
Chez les dicotylées ( voir plus loin) les stomates sont généralement
plus abondants à la face inférieure de la feuille ; chez les monocotylées, il y a équivalence numérique entre les deux faces foliaires : pourquoi ?3° Les hydathodes,
aussi appelés stomates aquifères servent à l'émission d'eau à l'état
liquide « et des solutés minéraux et organiques (phénomène de
guttation). Ils n'existent que sur les bords des feuilles, dans le
prolongement des nervures et leur nombre est beaucoup plus faible que
celui des autres stomates.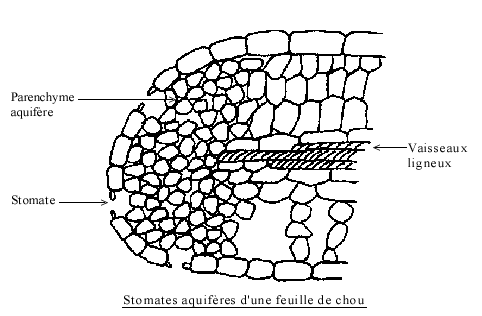 Un Un
stomate aquifère est constitué comme un stomate aérifère. Toutefois,
l'ostiole ne communique pas avec une chambre sous stomatique: à la place
de celle-ci, il y a un massif de petites cellules dépourvues de
chloroplastes, le tissu aquifère. A sa base se terminent les nervures.
Les cellules stomatiques sont dépourvues de chlorophylle: l'ostiole
reste donc toujours largement ouvert.4° Les lenticelles remplacent les stomates en structure secondaire ( voir plus loin).
5° Les papilles et poils épidermiques sont des extensions de cellules
épidermiques , ils sont de forme variable ( crochets, étoilés…) ou sont
parfois de simples papilles. Papilles et poils épidermiques sont à
considérer comme moyens supplémentaires de réduire l’intensité des
échanges gazeux ( transpiration).On distingue :- les poils unicellulaires : prolongement d'une cellule épidermique.- les poils pluricellulaires (formés de plusieurs cellules) qui peuvent être ramifiés
Les plantes qui doivent lutter contre une transpiration excessive , ou
les plantes qui vivent en milieux très secs ( rochers, vieux murs…)
peuvent avoir un revêtement épidermique velu ( léontondon, pissenlit…).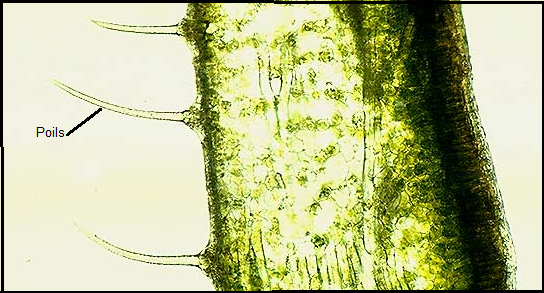 6° 6°
Les poils spécialisés sont par exemple des poils urticants, sécréteurs.
Ils sont unicellulaires ou non. La sécrétion est parfois localisée dans
la cellule terminale, ou parfois dans un décollement entre la membrane
externe ou le cytoplasme. 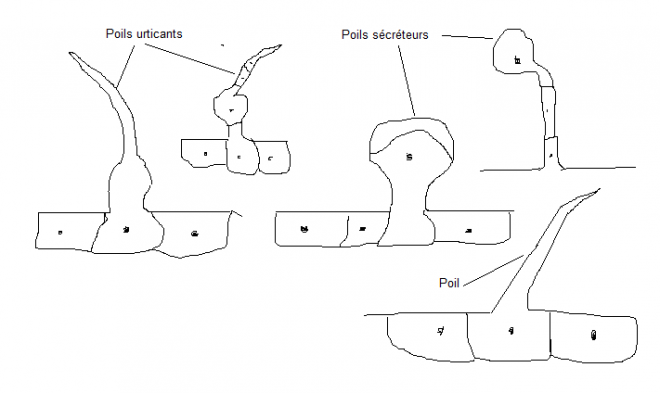 les les
orties sont connuespour leurs poils urticants . Ces poils sont nombreux
sur la tige et les nervures des feuilles. Chaque poil est constitué par
une partie basale, enflée, sécrétrice, et par une partie terminale, qui
est une cellule unique ; cette cellule a une pointe siliceuse, des
parois latérales incrustées de calcaire et une partie proximale gonflée
du liquide irritant, riche en acides. Les poils sont dirigés vers les
extrémités de la plante, que l'on peut donc caresser de bas en haut et
non à rebrousse-poil.Schéma : exemple d’adaptation à la sécheresse, stomates logés dans une crypte pourvue de poils épidermiques.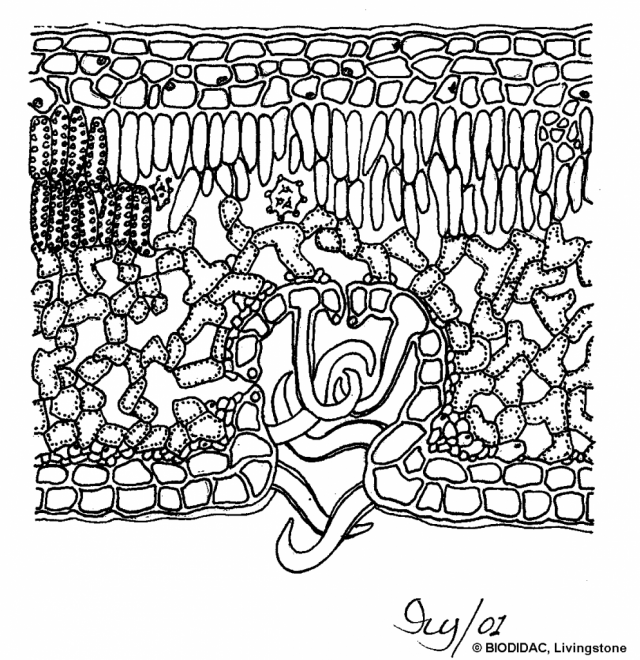 7° 7°
Le tissus subérisé ou liège est constitué de cellules imprégnée de
subérine mortes. Les cellules en sont rétrécies, ce tissus est destiné à
remplacer les épidermes défaillants ( par exemple, les épidermes des
tiges ligneuses).La subérine est un composé polyphénolique. 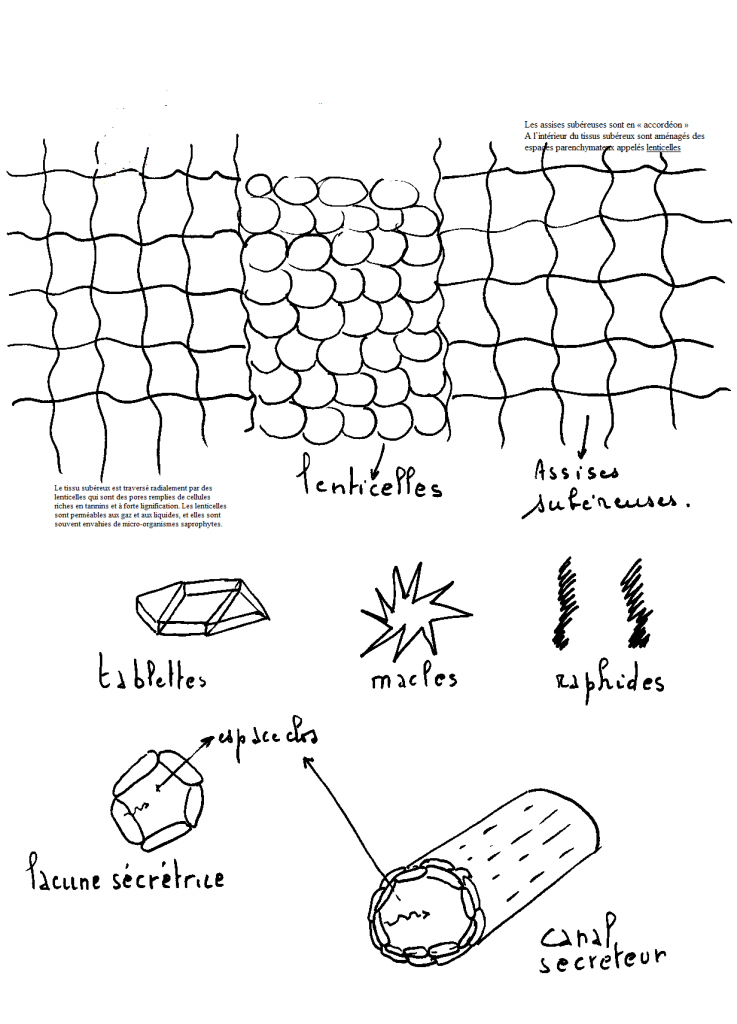 IV.8 LE TISSUS DE SECRETIONCe IV.8 LE TISSUS DE SECRETIONCe
genre de tissus résulte d’une activité cytoplasmique qui livre des
substances diverses telles que parfums, résines, latex, gommes,
cristaux. . .Les
substances élaborées sont parfois considérées comme des substances de
luxe, en ce sens qu’elles ne seraient pas nécessaires aux plantes qui
les produisent, du moins directement ??Ces
substances sont très variables et produites par le cytoplasme, elles
demeurent dans les cellules où elles ont été synthétisées ou bien
migrent à l’extérieur de celles-ci . On distingue :1° Les lacunes sécrétrices isolées qui peuvent être de plusieurs types, cellules epidermiques ( ortie, thym, houblon. . .), cellulesparenchymateuses , Laurier, oseille.Les
matières de sécrétion s’accumulent dans la vacuole sous forme de
cristaux ( macles, raphides, tablettes, ex : Oxalate de Calcium) ousous forme de produits non cristallisés ( parfums, huiles, alcaloïdes, résines.. . ).2° Des lacunes secrétrices : formation de grands méats par confluence , les cellules qui entourent la lacune sont sécrétrices et déversent
leur contenu dans la lacune qui sert de contenant . Ex : lacunes
sécrétriuces des orangers, des citronniers ( dans le zeste)
d’eucalyptus.3° Les canaux sécréteurs : proviennent de longs méats superposés. La matière de sécrétion est éliminée de la cellule sécrétrice et déversée dans l’espace ( canal ) situé entre les cellules sécrétrices.Autour
des cellules sécrétrices, existent selon le cas une ou plusieurs
assises fibreuses qui évitent les écrasements . Les substances sécrétées
sont selon le cas des résines ( « résineux »), des gommes ( cerisier), des mucilages etc…4° Des tubes à latex ou tubes laticifères : Il
s’agit, ici, contrairement aux cas précédents d’espaces
intracellulaires, de tubes plus ou moins ramifiés parcourant l’entièreté
de la plante, de structure symplastique et entourés de parenchyme . Le
latex est une substance laiteuse ou jaunâtre au centre du symplaste de
composition complexe ( huiles, glucides, alcaloïdes, protéines, tanins ,terpènes ).Et aussi des grains d’amidon de forme spéciale ( haltères). Exemple : Gutta percha ( Palaquium ) Chewing-gum ( Achras sapota), Caoutchouc ( latex de l’Hevea ), Opium ( Pavot). Les tubes laticifères contiennent un cytoplasme pariétal multinucléé et une grande vacuole centrale contenant le latex. Origine des tubes laticifères a) Une seule cellule qui grandit sans se cloisonner Hevea, Euphorbe, Ficus.
b) Plusieurs cellules qui deviennent un symplaste par résorption de
leur membrane transversale : Pissenlit, Salsifis, Pavot, Laitue.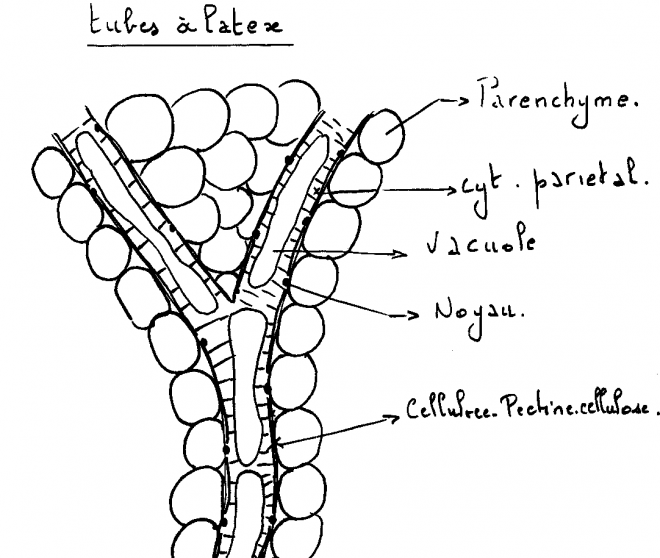 5° DES NECTAIRES 5° DES NECTAIRES
Ce sont des tissus situés sur certains réceptacles floraux et
sécrétant un exudat ( nectar, riche en sucres), les nectaires sont
souvent en rapport avec les conducteurs de la sève organique. Ils
favorisent la pollinisation à l’intervention d’insectes butineurs,
d’oiseaux, etc. . . | |
|

