| | | COURS DE NUTRITION ANIMALE DEUXIEME ANNEE (Par DR MEZIANE TOUFIK) |  |
|
+6Dakhi_55 laadjal mazagan merouane ZOOHVET amira 10 participants | | Auteur | Message |
|---|
amira

 Messages : 72 Messages : 72
 Points : 26 Points : 26
| Sujet: COURS DE NUTRITION ANIMALE DEUXIEME ANNEE (Par DR MEZIANE TOUFIK) 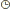 Jeu 20 Aoû 2009 - 11:57 Jeu 20 Aoû 2009 - 11:57 | |
| COMPOSITION CHIMIQUE DES ALIMENTS DU BETAIL I- ETUDE DES CONSTITUANTS DES ALIMENTS DES RUMINANTSLes herbivores et plus spécialement les ruminants représentés par l’importance des bovins , ovins et caprins , occupent une place prépondérante chez les animaux domestiques utilisés à des fins de production .Ils possèdent la particularité de transformer les végétaux non utilisables par le reste du règne animal en produits de grande valeur nutritionnelle pour l’être humain , telles que les protéines contenues dans la viande et le lait. Les ruminants sont les seuls à pouvoir valoriser les constituants cellulosiques des aliments d’origine végétale .Les ruminants domestiques tirent 90 à 95 % de leur nourriture de l’appareil végétatif aérien des plantes herbacées , des plantes vivrières après leur récolte et des arbustes . Ces fourrages sont d’une extraordinaire diversité dans leur nature botanique et leurs caractéristiques morphologiques anatomiques et physico-chimiques qui , toutes , agissent sur leur ingestibilité , leur dégradation dans le rumen et leur digestibilité.
I.1- LES CONSTITUANTS GLUCIDIQUESL’analyse chimique est beaucoup plus simple et reproductible que la détermination de la composition morphologique. Boussingault au 19éme siècle faisait de l’azote le critère principal de la valeur nutritive. Vers 1860 , Henneberg et Stohmann retinrent le principe du dosage de la cellulose brute par une double hydrolyse acide puis alcaline (Jarrige et al., 1995a). Une plante ne se décrit pas complètement à des fins nutritionnelles par un bulletin d’analyse . C’est d’abord un édifice d’organes , chacun constitué de tissus , dont la désintégration physique par la mastication est un préalable à la dégradation chimique et joue un rôle primordial dans la quantité ingérée . I.1.1- Les glucides cytoplasmiques Ce sont essentiellement des glucides hydrosolubles dont la digestibilité est totale .Le glucose , le fructose et le saccharose sont prédominants et représentent environ de 3 à 8% de la M.S. Cependant , il existe aussi des fructosanes qui s’accumulent à la base des tiges des graminées. La teneur des plantes fourragères reste en général assez faible .La concentration maximum est atteinte un peu avant le début de l’épiaison chez les graminées et un peu avant le bourgeonnement chez les légumineuses. L’amidon est absent généralement dans les fourrages sauf dans certaines légumineuses où des teneurs allant de 0.5 à 3 % de la MS ont été observées. I.1.2- Les glucides pariétaux Dans cette famille de glucides , il faut distinguer les polyosides proprement dits et les constituants pariétaux qui leur sont associés. I.1.2.1- Les polyosides sont formés de trois groupes : la cellulose , les hemicelluloses et les substances pectiques I.1.2.1.1 La cellulose : c’est un glucane formé de longues chaînes de molécules glucose aux environs de 1000 dans les plantes fourragères qui sont unies par des liaisons osidiques de type ß (1-4). Jarrige et al., 1995). La liaison osidique ß, contrairement à la liaison , est résistante à l’attaque des enzymes du suc digestif mais qui peuvent être attaquée par les enzymes des bactéries de l’appareil digestif des ruminants. Ces chaînes sont disposées parallèlement et associées en fibrilles puis en en fibres .Cette structure spécifique et sa configuration ß lui confère une certaine résistance aux enzymes et ce qui explique en partie sa digestibilité inférieure à celle des glucides pariétaux. (ITEB- INRAP 1984 , Jarrige et al., 1995a) La cellulose est le constituant principal des tissus de soutien (le collenchyme et le sclérenchyme ) et d’une partie des tissus de conduction (le xylème ). Dans les fourrages , elle représente de 40 à 45 % de l’ensemble des parois et par rapport à la M.S totale de la plante , la teneur varie de 15 à 40 % selon l’espèce et surtout selon l’âge de la plante. Cette proportion augmente avec l’âge de la plante et c’est ce qui explique en partie la diminution de la digestibilité lorsque la plante vieillit ( Jarrige. 1981, ITEB- INRA 1984). I.1.2.1.2- Les hémicelluloses : ils sont composés essentiellement de pentoses ,xylose en particulier ,de quelques hexoses et d’acides uroniques. Leur teneur varie de 12 à 25 % de la M.S des fourrages . Plus la plante vieillit ,plus la teneur en hémicelluloses augmente .Leur digestibilité est un peu inférieure à celle de la cellulose en raison de leur imprégnation par la lignine (Jarrige, 1981). I.1.2.1.3- Les substances pectiques qui sont des dérivés de l’acide galacturonique. On les retrouve dans les lamelles moyennes des cellules. Elles ont une teneur avoisinant les 2 % de la M.S des graminées et ayant une digestibilité très élevée et proche de celle des glucides cytoplasmiques (ITEB- INRAP, 1984). I.2- LES CONSTITUANTS NON GLUCIDIQUES Le seul constituant intéressant est représenté par la lignine qui est une substance complexe formée de plusieurs alcools. La lignine incruste progressivement les fibres polyosidiques des tissus de soutien et de conduction. Sa teneur varie de 2 % dans l’herbe jeune à 12 –13 % de la M.S dans la paille. La lignine est pratiquement indigestible , elle protège une partie des polyosides membranaires ,qu’elle incruste, de la dégradation microbienne .De ce fait la lignine est le facteur principal limitant de la digestibilité des aliments .I.2.1- LES CONSTITUANTS AZOTES (ou matières azotées totales = protéines brutes ) I.2.1.1- Les matières azotées protidiques (MAP): Elles sont localisées dans les cellules chlorophylliennes et elles donnent par hydrolyse des acides aminés. Elles sont constituées de protéines , de peptides et des acides aminés libres. I.2.1.2- Les matières azotées non protidiques (MANP ): Elles sont localisées dans les vacuoles des cellules végétales , elles ne donnent pas à l’hydrolyse des acides aminés .Ce sont les amines , les amides (telle que l’urée,…), les formes azotées simples (NO2-, NO³-, NH4+…), les bases azotées (formes cycliques constituants des acides nucléiques ). Dans les fourrages verts, elles représentent 15 à 35 % des matières azotées totales, cette proportion est plus élevée dans les tiges que dans les feuilles .Les légumineuses sont plus riches que les graminées. Les matières azotées non protidiques des foins récoltées dans de bonnes conditions ont en général une proportion comprise entre 30 et 40 % des matières azotées totales (ITEB-INRAP, 1984). I.2.2- LES CONSTITUANTS LIPIDIQUES . Dans les plantes fourragères ,on trouve des galactolipides qui sont des glycérides associés à du galactose .Ils sont localisés dans les chloroplastes , riches en acides gras non saturées et en particulier l’acide linoléique .On rencontre également des cérides (qui sont des alcools à poids moléculaire élevé + des acide gras ) dans la cuticule des feuilles où ils constituent la substance principale de la cutine qui est indigestible . Les lipides représentent une très faible fraction de la M.S des fourrages (2 à 5 %), ce qui explique le peu d’intérêt qu’il leur est accordé le plus souvent (Jarrige 1995a). Tableau n° 1 : Les différents constituants des aliments (ITEB- INRAP, 1984) 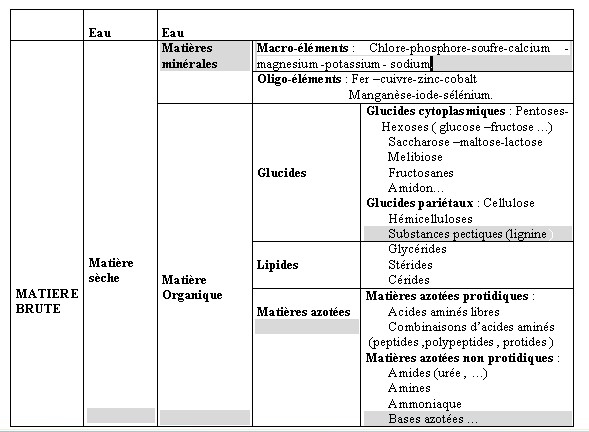 Tableau n° 2 : Les principaux glucides cytoplasmiques et constituants pariétaux ( Jarrige, 1981) 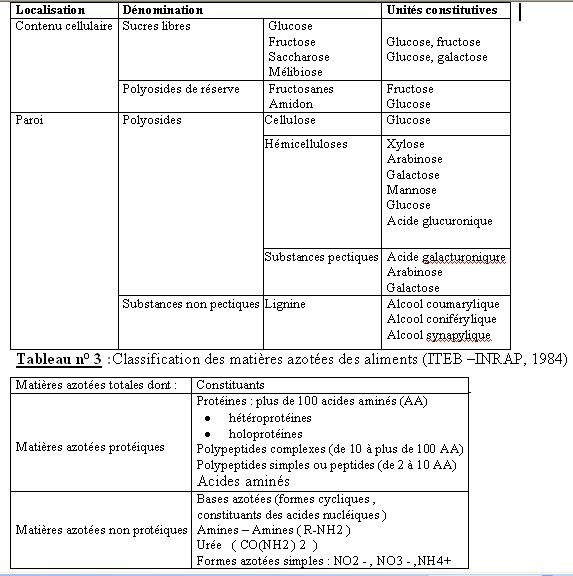 I.2.3- LES MINERAUX : I.2.3- LES MINERAUX :La composition minérale d’un fourrage résulte de l’action de plusieurs facteurs comme le stade de végétation de la plante , sa famille botanique et les conditions de milieu et d’exploitation (comme les épandages ou les fertilisants ). Comme tous les mammifères , les ovins doivent trouver dans leur régime alimentaire tous les éléments minéraux indispensables en quantités suffisantes . Compte tenu des risques d’insuffisance d’apport en éléments minéraux des rations à base de fourrages des ruminants , l’attention des nutritionnistes est attirée sur les éléments minéraux suivants: P , Ca , Na , Mg , S ( pour la laine ), Zn ,Cu ,Co (Gueguen et al., 1978 ). Avant d’aborder ,le métabolisme minéral et les besoins des animaux , il convient d’évaluer l’apport alimentaire qui recouvre la composition minérale des aliments des ruminants et la capacité d’absorption de l’animal. Les matières minérales totales (ou cendres brutes ) représentent de 8 à 15 % de la matière sèche ( MS) des fourrages (Meschy et al., 1995). 1.2.3.1- Les macro-éléments minéraux : se présentent dans le végétal sous des formes chimiques variées : * Le potassium et sodium sont presque totalement ionisés . * Les phosphates sont sous des formes multiples ; P inorganique ; Phosphates estérifiés ou non ; phytates dans les graines . * Le calcium est sous forme soluble ,partiellement soluble (phosphates) ou sous forme d’oxalates insolubles , et enfin une fraction de calcium peut également être liée aux protéines et aux pectines * Le magnésium dont 50 % sont sous forme soluble , environ 10 % associés au complexe chlorophyllien , et une partie non négligeable est complexée aux acides organiques et à la lignine (Meschy et al., 1995 ). Tableau n° 4 : Plages de variations des teneurs minérales dans les fourrages ( Little,1982 cité par Meschy et al., 1995 Macro-éléments Teneur en g /kg de MS P 0.2 à 7 Ca 0.4 à 41 Mg 0.3 à 10 Na 0.01 à 21 K 10 à 60 S 0.5 à 4 1.2.3.2- Les oligo-éléments Les plantes fourragères peuvent souffrir de carences en oligo-éléments avec une production diminuée dues essentiellement aux carences rencontrées principalement sur les sols .Selon Grace et al.,1991, l’usage de fertilisants peut altérer la concentration des oligo-éléments dans les plantes en augmentant la teneur en Mo et Se et en diminuant celles du Co, Mn, Fe et Zn . Tableau n° 5 : Teneurs en oligo-éléments de quelques grains et fourrages (en mg /kg de MS ou ppm) (INRA. 1978 ; Chapuis. 1991). 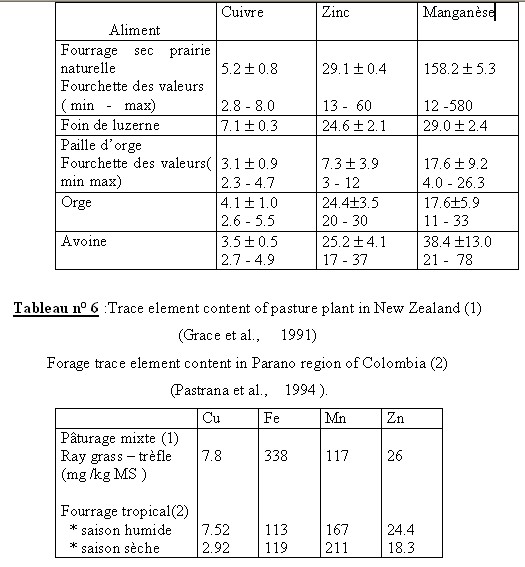 Tableau n ° 7 : Répartition des teneurs en oligo-éléments des foins de prairie naturelle(en mg /kg de MS ) (Lamand et al., 1981) 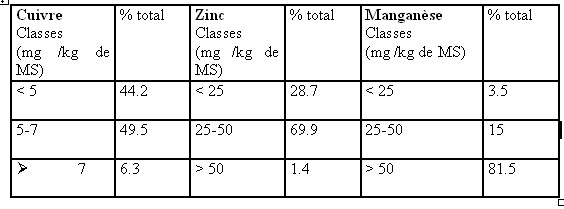 | |
| | | | amira
Messages : 72
Points : 26
| | Sujet: Re: COURS DE NUTRITION ANIMALE DEUXIEME ANNEE (Par DR MEZIANE TOUFIK) Jeu 20 Aoû 2009 - 12:20 | |
| II - UTILISATION DIGESTIVE ET METABOLIQUE DES ALIMENTS
II .1- Rappel anatomique du tube digestif des ruminants L’appareil digestif des ruminants est caractérisé par un estomac très différencié à plusieurs cavités comprenant quatre parties nettement distinctes extérieurement. Ce sont successivement le rumen (ou panse), le réseau (ou réticulum) et le feuillet (ou omasum).Ces trois premières représentent les pré-estomacs et sont placées avant l’estomac appelé la caillette .Toutes les parties de l’estomac des ruminants dérivent d’une ébauche simple et il faut les considérer comme le résultat d’une différenciation spécifique et d’une adaptation à la nature particulière de l’alimentation. Les ruminants sont des herbivores qui peuvent digérer les parois des tissus végétaux non lignifiés dont la cellulose est le constituant principal. II.1.1- le rumen ou la panse ,c’est le plus volumineux, il renferme de 70 à 75 % du contenu du tube digestif et représente de 50 à 60 % de son volume . Son volume et celui du réseau sont d’environ 18 litres. II.1.2- le réseau (ou réticulum ) qui doit son nom à sa muqueuse réticulée et parsemée de papilles absorbantes , joue un rôle central dans la circulation des particules .Les particules qui franchissent l’orifice réticulo-omasal doivent avoir une taille moyenne inférieure ou égale à 1 mm. De ce fait , les aliments solides sont donc séquestrés tant qu’ils n’ont pas atteints cette taille minimale .( Pearce 1967 , Ulyatt 1983 , Reid 1984 , Ulyatt et al., 1985 ) cités par Sutherland, 1986 ; Thivend et al., 1985. II.1.3- le feuillet (ou omasum ) : il doit son nom de feuillet au fait qu’il est presque entièrement occupé par des lames parallèles , de hauteurs inégales ,disposées dans le sens du transit alimentaire . D’un volume égal à 0,5 l , c’est un organe ovoïde chez le mouton à l’intérieur duquel on trouve de très nombreuses lames recouvertes d’un épithélium kératinisé possédant également des papilles. Il communique en aval avec la caillette par un orifice large et dilatable II.1.4- La caillette est le seul réservoir sécrétoire de l’estomac des ruminants .Sa cavité est tapissée par une muqueuse glandulaire , analogue à celle des monogastriques toujours recouverte d’une couche de mucus .Les fonctions digestives de la caillette des ruminants sont analogues à celles de l’estomac des mammifères mono-gastriques .Elle a un volume inférieur à 2 litres .L’ensemble de ces réservoirs ont une capacité de 12 à 20 litres chez le mouton . II.1.5- L’ intestin Il est divisé en deux parties : • L’intestin grêle est très long. Il comprend le duodénum ( 0,6 à 1,2 m )avec son anse duodénale qui reçoit les sécrétions biliaires et pancréatiques et l’ensemble jéjunum-iléon (17,5 à 34,0 m )(Nickel et al., 1987 ).Sa structure est identique à celle de l’être humain .Les mécanismes de la digestion et de l’absorption dans l’intestin grêle sont les mêmes que chez les mono-gastriques . • Le gros intestin comprend le cæcum , le colon sigmoïde , le colon spiral , le colon flottant et le rectum .Le gros intestin ne secrète pas de sucs digestifs. II .2- DIGESTION DANS LE RUMEN – RESEAU La digestion est réalisée grâce à des enzymes cellulolytiques que les micro-organismes du rumen , du réseau et du gros intestin peuvent sécréter. La présence de la dégradation microbienne dans les pré-estomacs avant la digestion( chimique ) par les sécrétions digestives dans la caillette modifie fortement la digestion et l’utilisation des aliments par les ruminants par rapport aux mono-gastriques. La digestion chez les ruminants met en jeu des phénomènes mécaniques , des phénomènes fermentaires ou encore biologiques en relation avec la présence d’une flore (au niveau du rumen - réseau ) et enfin des sécrétions enzymatiques en relation avec les sécrétions digestives (dans la caillette et l’intestin grêle comme chez les mono-gastriques ). L’ingestion des aliments par les poly-gastriques (qui dure environ 8 heures par jour avec une mastication rapide au cours de la quelle les aliments s’entassent dans le rumen) est suivie par la rumination. La rumination est un état physiologique défini comme un phénomène cyclique qui se déroule en quatre étapes : Diagramme n° ( 1) : Cycle rumination –digestion ( Ulyatt, 1982 cité par Preston et al., 1987) 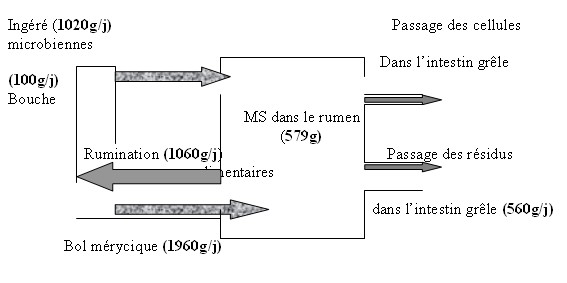 Ce diagramme montre les flux de la matière sèche à travers le rumen du mouton alimenté avec 1020g de foin haché de luzerne dans un intervalle de 24 heures a- La régurgitation du bol alimentaire comportant deux phases : * Une phase d’aspiration œsophagienne au cours de laquelle le réseau se contracte ce qui amène le contenu du rumen au niveau du cardia. Le cardia s’ouvre et l’animal inspire fortement créant une dépression intra-thoracique. Cette dernière permet au contenu du rumen de remonter dans l’œsophage . * Une phase d’expulsion vers la bouche, due à une onde antipéristaltique accompagnée d’une expiration profonde . b- La déglutition de la partie liquide servant de support. c- La mastication mérycique au cours de laquelle les mouvements sont lents et la salivation est abondante . d- La phase de repos . Il existe 6 à 8 périodes de rumination par jour ,en moyenne de 40 à 50 minutes chacune. La rumination est indispensable car elle fragmente les aliments et facilite l’attaque par les micro-organismes du rumen. La complexité anatomique de ces réservoirs est associée à la présence d’une population dense et stable de micro-organismes qui vivent en symbiose avec l’hôte et qui jouent un rôle dans la digestion et la nutrition de l’animal. La mise en place et le maintien de cette population microbienne sont dus au fait qu’il existe une séparation physique entre la zone de sécrétion acide (estomac ) et le reste des préestomacs où la digestion microbienne peut avoir lieu en permanence (Jarrige 1995 b). | |
| | | | amira
Messages : 72
Points : 26
| | Sujet: Re: COURS DE NUTRITION ANIMALE DEUXIEME ANNEE (Par DR MEZIANE TOUFIK) Jeu 20 Aoû 2009 - 12:21 | |
| II .2.1- Le contenu du milieu ruminal Le contenu du milieu ruminal est relativement constant , il se caractérise par : - une concentration élevée en eau 85 à 90 % ; - une température constante de 39 à 40° C ; - un potentiel d’oxydo-réduction variant entre de – 250 à - 400 mV(milieu fortement anaérorobie ) ; - un pH généralement compris entre 6 et 7 qui est tamponné par l’apport régulier de grandes quantités de bicarbonates et de phosphates contenus dans la salive ; - une pression osmotique constante proche de celle du sang ; - un apport régulier de nutriments et d’eau fournis à la fois par l’ingestion des aliments et par la rumination ; - une élimination continue des produits du métabolisme , soit par absorption à travers la paroi du rumen ( acides gras volatils , ammoniac ) , soit par passage dans la partie postérieure du tube digestif (résidus alimentaires, cellules microbiennes ), soit par éructation ( méthane ,gaz carbonique ) ; - une relative constance de l’atmosphère gazeuse située au niveau du sac dorsal (gaz carbonique : 60 –70 % , méthane : 30 – 40 % ) ; - un brassage permanent assuré par les contractions périodiques de la paroi et par la rumination . Le rumen est donc un milieu particulièrement bien adapté au développement d’une population microbienne anaérobie dont les principaux constituants sont les bactéries et les protozoaires .(Thivend et al., 1985 , ITEB-INRAP 1984 ; Russel 1986 , Preston et al., 1987 ). II.2.1.1- Les protozoaires .Ce sont principalement des ciliés dont la taille varie de 20 à 200µ mais on observe aussi des flagellés en nombre réduit . Le nombre total des protozoaires se situe aux environs de 100.000 à 1000000 ( ITEB-INRAP, 1984 ) ou 2 à 5000000 par ml de jus de rumen (Thivend et al., 1985). Les protozoaires ciliés sont capables de transformer un grand nombre de constituants alimentaires et bactériens en métabolites et en composés cellulaires qui seront ensuite utilisés par l’animal hôte .Ils appartiennent à deux groupes , les Holotriches et les Entodiniomorphes. Ces protozoaires sont capables pour la plupart d’entre eux de dégrader la cellulose, les hémicelluloses, les pectines pour les entodiniomorphes et les sucres solubles pour les Holotriches. Les ciliés sont également protéolytiques, ils utilisent aussi les acides aminés (Coleman 1980 cité par Thivend et al.,1985 )mais leurs besoins azotés sont en grande partie couverts par l’ingestion des bactéries .Les protozoaires sont généralement libres dans le liquide du rumen mais certains se fixent également aux particules alimentaires ( Beauchop et al., 1976 cités par Thivend et al., 1985 ). Ils sont très sensibles aux conditions régnant dans le rumen et aux caractéristiques de la ration. Avec des rations riches en glucides solubles ou en amidon, ils peuvent représenter au maximum 50 % de la biomasse du rumen, et ils régressent avec les rations très pauvres en constituants solubles. Ils peuvent disparaître avec des rations créant des pH bas du rumen (< 5,5 )aliments concentrés distribués à volonté et / ou séjournant pendant un temps très court dans le rumen (fourrages broyés ). II.2.1.2- Les bactéries La population bactérienne du rumen est comprise entre 8 et 10 milliards cellules par ml de contenu ruminal. Elle constitue environ 50 % de la biomasse microbienne et représente la catégorie des micro-organismes la plus complexe et la plus importante. Elle est composée essentiellement de bactéries anaérobies strictes non sporulées et elle est caractérisée par sa très grande diversité .En 1959 on en dénombrait 39 genres et 63 espèces et actuellement plus de 200 espèces bactériennes ont été isolées du rumen mais seulement une trentaine d’entre elles peuvent être considérées comme des bactéries authentiques du rumen alors que les autres sont apportées par les aliments et sont présentes d’une manière transitoire (Russel et al., 1981cités par Thivend et al.,1985 ; Russell 1986 ). Les bactéries du rumen sont généralement classées en fonction des substrats qu’elles sont capables de fermenter ou de dégrader; on peut citer les bactéries cellulolytiques, pectinolytiques, amylolytiques , uréolytiques … Les fonctions de plusieurs espèces se recouvrent largement ce qui contribue à la stabilité de l’ecosystème. Cependant certaines d’entre elles sont plus spécialisées telle que Anaerovibrio lipolytica qui n’hydrolyse que les lipides et ne fermente que le glycérol ou bien les Veillonella qui utilisent principalement le lactate (Hungate 1966 cité par Jarrige 1995a), Vibrio succinogenes ne tire son énergie que de la réduction du fumarate en succinate (Thivend et al., 1985 ). De nombreuses espèces sont protéolytiques , plusieurs d’entre elles utilisent ou dégradent les acides aminés ou les peptides. (Russell 1986., Preston et al., 1987 ) Dans le rumen, les bactéries occupent trois biotopes distincts : elles peuvent être libres dans le liquide ruminal , ou attachées soit à la paroi interne du rumen, soit aux particules alimentaires alors que d’autres espèces vivent liées à la surface des protozoaires environ 1 à10% de la flore totale (Imai et Ogimoto , 1978 cités par Jarrige et al., 1995b.) Les bactéries attachées aux particules alimentaires représentent environ la moitié de la population bactérienne selon Minato et al.,1966 cités par Thivend et al.,(1985) , elles concernent principalement celles qui hydrolysent les polymères pariétaux. Les bactéries adhérentes à la paroi du rumen appelée flore épimurale sont caractérisées par leur forte activité protéolytique (Dinsdale et al., 1980 cités par Thivend et al., 1985) et uréolytique (Mc Cowan et al,1980 cités par Thivend et al., 1985 ; Wallace 1986). Ses bactéries ont un rôle primordial parmi lesquels , on citera : • L’hydrolyse de l’urée qui diffuse à travers la paroi du rumen . • La dégradation des cellules épithéliales fortement kératinisées provenant de la desquamation de la muqueuse du rumen. • L’élimination de l’oxygène qui diffuse à travers la paroi du rumen depuis la voie sanguine([ Dinsdale et al., 1980 , Mc Cowan et al., 1980] cités par Thivend et al., 1985).Ces bactéries sont aussi très riches en phosphatase alcaline ( Fay et al.,1979 cités par Thivend et al., 1985). Lorsqu’un animal consomme un régime alimentaire uniforme, il s’établit dans son rumen , après une période d’adaptation , une population microbienne de composition relativement constante , qu’on peut appeler un faciès microbien. L’état d’équilibre est assez fragile et conditionne l’efficacité de la digestion. II.2.1.3- Autres micro-organismes
II.2.1.3.1- Les champignons L’existence des champignons anaérobies dans le rumen a été démontrée par Orpin en 1977 et Bauchop 1979, Joblin 1981 cités par Thivend et al., 1985. Bauchop a indiqué que ces champignons étaient liés à la fraction solide du contenu du rumen et que l’ importance de leur effectif dépendait de la présence de fourrages grossiers dans la ration .Ils existent chez les animaux nourris avec des aliments succulents ou qui pâturent une herbe jeune (Jouany, 1981 et Czerkawski., 1984). La plupart de ces micro-organismes apportés par l’aliment seraient simplement en transit dans le rumen Certains Phytomycètes anaérobies stricts joueraient un rôle non négligeable dans la dégradation des polyosides pariétaux ( Bauchop 1981 et Orpin 1984 cités par Thivend et al.,1985 ; Jarrige et al, 1995b) et probablement dans la dégradation du méthane (Preston et al., 1987 ). II.2.1.3.2- Mycoplasmes , virus et bactériophages Ils sont présents dans le rumen mais leurs rôles restent à élucider surtout pour les mycoplasmes .Selon Thivend et al., 1985 , les bactériophages tempérés et virulents peuvent jouer un rôle dans l’écologie du rumen .A la suite d’une infection par un phage virulent , une espèce bactérienne peut devenir sous dominante Son action pourra cependant être compensée par la prolifération d’une espèce non sensible , capable d’occuper la même niche écologique II.2.2- La dégradation des glucides Grâce à un extraordinaire équipement enzymatique, la population microbienne du rumen et du réseau hydrolyse tous les glucides en oses (hexoses ou pentoses ). Les glucides solubles sont hydrolysés de manière très rapide et en totalité (amidon des céréales est dégradé à 90 – 95 % dans le rumen par une amylase bactérienne ). La population microbienne dégrade aussi les glucides des parois (cellulose, hémicelluloses et pectines ) en oses qui sont ensuite fermentés en anaérobiose selon des voies bien connues (voir schéma du métabolisme des glucides pariétaux dans le rumen ). Cette dégradation par des cellulases que seules les bactéries sont capables de synthétiser, est lente et partielle ( de 80 à 90 % pour les fractions pariétales pour un aliment très peu lignifié comme l’herbe jeune , à 40 50 % pour une plante riche en lignine comme la paille ). Cette hydrolyse libère du cellobiose (2 ß glucose ), du glucose et des pentoses. La population microbienne du rumen –réseau tire de la fermentation des oses provenant de l’hydrolyse des glucides pariétaux, l’énergie (ATP ) et le carbone qui lui sont nécessaires pour son entretien ,sa croissance et sa prolifération. Les produits de cette fermentation sont un mélange d’acides organiques à courte chaîne, dits acides gras volatils, essentiellement acétique, propionique et butyrique ,et des gaz (gaz carbonique et méthane ).Les produits intermédiaires (acides succiniques et lactique ) sont généralement utilisés par les microbes au fur et à mesure de leur formation .Le ruminant trouve la majeure partie de l’énergie dont il a besoin dans les acides gras volatils (AGV ) issus de la dégradation des glucides .Ils peuvent lui fournir de 65 à 75 % de l’énergie absorbée . Avec les régimes habituels à base de fourrage utilisés par les ruminants , les proportions relatives des AGV sont les suivantes (en % molécules ) - Acide acétique (C 2) 60 à 70 % - Acide propionique ( C3 ) 15 à 20 % - Acide butyrique ( C4 ) 10 à 15 % - Autres AGV 2 à 5 % Les gaz formés au cours des processus fermentaires ,CO2 et CH4 représentent des pertes énergétiques pour les ruminants et sont rejetés par éructation . La production de ces gaz dépend de l’intensité des fermentations microbiennes ; elle augmente ainsi avec la digestibilité de la matière organique de la ration et diminue en fonction des facteurs qui limitent le temps de séjour des aliments dans le rumen ( comme le broyage ,l’agglomération , l’action des agents chimiques ou biologiques). La diminution du temps de séjour , modifie le temps de contact enzyme-substrat ,accroît la surface pariétale accessible aux enzymes et réduit le nombre et la force des glucides –lignine qui limitent l’action des micro-organismes (Czerkawski 1984 ; Thivend et al., 1985 ; Jarrige et al., 1995b). Schéma n ° 1 : Le métabolisme des glucides pariétaux dans le rumen (d’après Van Soest , 1982 cités par Thivend et al.1985) 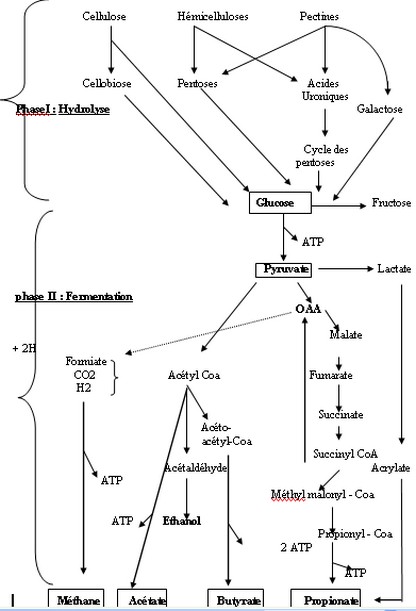 à suivrr | |
| | | | ZOOHVET
Messages : 79
Points : 70
| | Sujet: Re: COURS DE NUTRITION ANIMALE DEUXIEME ANNEE (Par DR MEZIANE TOUFIK) Jeu 20 Aoû 2009 - 12:48 | |
| 1000 merciiiiii Dr amira c'est très intéressent | |
| | | | amira
Messages : 72
Points : 26
| | Sujet: Re: COURS DE NUTRITION ANIMALE DEUXIEME ANNEE (Par DR MEZIANE TOUFIK) Jeu 20 Aoû 2009 - 13:13 | |
| II.2.3- La dégradation des matières azotées : Les matières azotées alimentaires subissent dans le rumen une dégradation plus ou moins intense et rapide dont l’ammoniac (NH3) est le produit terminal le plus important .Cette dégradation en ammoniac (protéolyse microbienne ) est rapide et totale pour les constituants non protidiques (urée , amides …) ainsi que pour les constituants protidiques simples (acides aminés libres , peptides et polypeptides ). Les substrats carbonés et l’ammoniac peuvent ensuite être pour la synthèse des matières azotées de certaines bactéries (cellulolytiques ) ce qui correspond à la phase de protéosynthèse microbienne . Dans la mesure où il n’est pas utilisé par les micro-organismes pour cette synthèse de matières azotées microbiennes , la majeure partie de l’ammoniac restant est absorbé au niveau de la paroi du rumen , véhiculé au foie où il est transformée en urée .Cette urée est en partie recyclée dans la salive ou par diffusion à travers la paroi de tout le tube digestif , et en partie éliminée par l’urine et donc perdue . L’uréogénèse à partir de l’ammoniac nécessite beaucoup d’énergie .Elle est estimée à 4 ATP / moles d’urée produite ( Remesy et Demigne, 1981 ). Selon Milton et Ternouth (1984) cités par Komisarczuk (1985), l’absorption d’ammoniac est conditionnée par sa concentration dans le rumen ( 50 à 80 mg / 100 ml de jus de rumen ) et par le pH du rumen (un pH élevé conduit à une absorption rapide , un pH bas à une absorption lente . Les bactéries qui vivent sous des concentrations basses d’ammoniac fixent l’ammoniac à deux étapes de la glutamine synthétase et la glutamate synthase au cours desquelles il y’a transfert de l’amide-N de la glutamine au 2-oxoglutarate et cette étape nécessite de l’ATP (Leng et al., 1987). La concentration en ammoniaque dans le rumen peut varier de 2 à 40 mmoles /l mais on estime à 4-5 mmoles /l la teneur nécessaire pour que la production de protéines bactériennes soit maximum (Thivend et al., 1985 ; Leng et al., 1984 ). Selon Vérité et al., 1987 , de 2/3 à 3/4 des fractions azotées des fourrages sont dégradés dans le rumen .Chez les moutons normalement alimentés , l’absorption est en moyenne de 4 à 5 g d’N 2 par jour sous forme d’NH 3 (Kolb, 1975) . Houpt , Decker et coll . cités par Kolb 1975 ont montré que chez les sujets à régime pauvre en protéines ,il n’y a guère que la moitié de l’urée du sang qui soit éliminée avec l’urine , le reste revient par diffusion dans la panse et peut servir à la synthèse de protides. Donc prés de 50 % de l’urée synthétisée par le foie suit un cycle rumino- hépatique et ne constitue pas un produit final du métabolisme mais une véritable réserve d’azote pour les ruminants . II.2.4- La dégradation des lipidesLes lipides ,en général faiblement présents dans la ration ,sont constitués d’acides gras en C18 non saturés, linolénique dans les fourrages ,linoléiques dans les grains Le contenu du rumen possède in vivo comme in vitro , une forte activité lipolytique vis à vis des glycérides d’acides gras supérieurs .La population microbienne hydrolyse les triglycérides en acides gras et glycérol .Le glycérol est fermenté en acides gras volatils et rejoint le circuit des glucides . Par ailleurs , la flore ruminale hydrogène les doubles liaisons des acides gras insaturés et incorpore une partie dans leurs lipides et les transforment en acide stéarique (C18 saturé ) qui sera digéré dans l’intestin (Soltner, 1994). Les acides gras à longue chaîne d’origine alimentaire sont fixés sur les particules alimentaires avec lesquelles ils passent dans le feuillet ou bien ils sont repris par les bactéries qui en élaborent leurs propres acides gras . II.2.5- Les éléments minéraux et la digestion microbienne au niveau du rumen-réseau.Comme tous les mammifères ,les ovins doivent trouver dans leur régime alimentaire tous les éléments minéraux indispensables en quantités suffisantes. Compte tenu des risques d’insuffisance d’apport en éléments minéraux des rations à base de fourrages des ruminants, l’attention des nutritionnistes est attirée sur les éléments minéraux suivants: P, Ca, Na, Mg, S ( pour la laine ), Zn ,Cu ,Co (Gueguen et al., 1978 ). Les micro-organismes du rumen , des quels dépend l’utilisation des fourrages ont des besoins minéraux propres , notamment en P ,S , Mg ,Zn ,Cu ,Co .Si ces besoins ne sont pas couverts, l’efficacité de la ration sera faible (Gueguen et al., 1988 ). Les plantes fourragères peuvent souffrir de carences en oligo-éléments avec une production diminuée dues essentiellement aux carences rencontrées principalement sur les sols .Ces carences sont responsables de la baisse des productions et de la reproduction des animaux qui y pâturent ( Pastrana et al., 1991). Les teneurs en oligo-éléments des micro-organismes du rumen sont généralement bien supérieures à celles des aliments que l’animal prend. Les parois des bactéries sont capables de fixer des oligo-éléments par des liaisons plus ou moins réversibles en milieu acide (Thivend et al., 1985 ). Certains oligo-éléments comme le fer(Fe ) ,le manganèse (Mn ), le zinc (Zn ), le cobalt (Co ), le molybdène (Mo) régulent de nombreuses activités enzymatiques bactériennes ou font partie de ces molécules organiques comme : * Cuivre : cytochrome oxydase , lysyl-oxydase ,tyrosinase ,céruloplasmine ,.. * Manganèse : pyruvate carboxylase * Zinc : anhydrase carbonique , aldolase , peptidase ,phosphatases alcalines , DNA et RNA polymérases ,deshydrogénases. *Certains minéraux majeurs (P et S) jouent un rôle essentiel dans l'optimisation des fermentations dans le rumen.( Komisarczuk, 1985 ; Komisarczuk et al., 1991) Certains d’entre eux rentrent dans la composition d’éléments cellulaires comme les ribosomes ou les membranes . Des études in vitro ont montré que l’activité cellulolytique est stimulée par des apports d’oligo-éléments , de même que la croissance des protozoaires en milieu continu peut être accrue par un apport de zinc ou de cobalt (Thivend et al., 1985) . II.2.6 - Devenir des corps microbiens La quantité de substances microbiennes formée est en moyenne proportionnelle à la quantité d’énergie disponible dans le rumen , ou à la quantité de matière organique digestible qui y disparaît , lorsque les apports de matières azotées fermentescibles , d’autres éléments nutritifs et de facteurs de croissance ne sont pas limitants . La majeure partie des corps bactériens formés passe dans le feuillet , en suspension dans le liquide ou fixée sur les résidus alimentaires .Un partie est détruite à l’intérieur du rumen par différents mécanismes , elle serait la plus importante dans le cas des protozoaires . II.2.7- Conclusion Il sort du rumen –réseau la majeure partie des corps bactériens et une partie plus faible des protozoaires. En ce qui concerne les constituants organiques des aliments , les quantités dégradées dans le rumen –réseau dépendent notamment de la nature de la ration . II.2.7.1- Cas des fourrages : sont dégradés dans le rumen –réseau :De 40 à 70 % des matières azotées , pratiquement la totalité des glucides solubles , la majeure partie des polyholosides membranaires des fourrages non broyés : * 90 à 95 % de la cellulose * 80 à 90 % des hémicelluloses :ces proportions pourraient être sensiblement plus faibles dans le cas de fourrages très lignifiés. Au total , 60 à 65 % de la matière organique digestible disparaît en moyenne dans le rumen – réseau . Remarque : le broyage des fourrages secs diminue le temps de séjour dans le rumen et l’activité cellulolytique de la population microbienne , il s’ensuit une diminution de la dégradation des membranes dans le rumen , une diminution de la quantité des AGV et de CH4 formés , une augmentation des protéines alimentaires non fermentées dans le rumen.(ITEB –INRAP, 1984 ; INRA 1995)
II.2.7.2- Cas des aliments concentrés : Environ 95 % de l’amidon de l’orge ,le blé et l’avoine sont dégradés dans le rumen Cependant, ce taux est nettement inférieur et aussi plus variable dans le cas de l’amidon de maïs et de sorgho (70 à 85 %) selon la quantité dans la ration .Cette teneur peut être augmentée par certains traitements hydro-thermiques .Les protéines des aliments concentrés sont dégradés en proportion très variable dans le rumen, elles sont de l’ordre de 30 à 90 % (ITEB –INRAP, 1984). II.2.7.3- Cas des rations mixtes Les interactions physiques entre les différents constituants de ce type de ration entraîne une diminution de la proportion de la matière organique digestible qui disparaît dans le rumen . Elle est plus variable pour les rations mixtes que pour les fourrages seuls .Cette diminution porte surtout sur les polyholosides membranaires par suite d’une diminution de la capacité cellulolytique de la population microbienne du rumen .Il sort du rumen réseau : * La majeure partie des corps bactériens * Une partie plus faible des protozoaires * 35 à 40 % de la matière organique digestible de la ration constituée surtout de matières azotées et de polyholosides membranaires à un moindre degré.(ITEB-INRAP, 1984). La population microbienne fournit à l’animal la majeure partie des substrats énergétiques (AGV) et des acides aminés , ainsi que des vitamines B en quantité suffisante (Thivend et al., 1985).  à suivre | |
| | | | amira
Messages : 72
Points : 26
| | Sujet: Re: COURS DE NUTRITION ANIMALE DEUXIEME ANNEE (Par DR MEZIANE TOUFIK) Ven 21 Aoû 2009 - 11:02 | |
| III- DIGESTION APRES LE RUMEN –RESEAU
III.1- Dans le feuillet : Longtemps considéré comme un lieu de passage et non d’absorption des nutriments. Le feuillet se remplit de façon cyclique par l’orifice réticulo-omasal qui laisse pénétrer les particules fines. Une partie des digesta s’écoule vers la caillette mais les deux tiers environ du flux digestif séjournent entre les lames du feuillet pendant une durée sans doute variable. La fraction ADF(Acid Detergent Fiber ou lignocellulose ) indigestible séjourne environ 2 heures dans le feuillet (Holtenius et al., 1989). Les digesta qui pénètrent dans l’omasum ont pour origine le contenu du plancher du réticulum et ont une teneur en matière sèche très faible de 3 à 8 % ( Moir, 1984 cité par Deswysen et al.,1995). Ce contenu réticulaire est renouvelé à chaque contraction par des digesta provenant du sac crânial du rumen ( Baumont et al., 1991 ). L’absorption des nutriments au niveau du feuillet fut étudiée par Engelhardt et Hauffe en 1975 cités par Deswysen (1995 ) qui ont démontré l’absorption de l’eau à ce niveau chez le mouton et la chèvre. Beaucoup d’auteurs ont montré que l’absorption omasale est réelle pour l’eau , les électrolytes et les AGV qui ont échappé à l’absorption du rumen et qui représentent environ 10 % de la production totale des AGV dans les pré-estomacs du mouton (Engelhardt et al., 1975 cités par Deswysen, 1995). En utilisant des marqueurs avant l’abattage des animaux Holtenius et al.,1989 , calculèrent une absorption omasale de l’eau séjournant dans l’espace interlamellaire de 15 % chez le mouton. Mais, en raison du passage direct d’une partie du liquide par le canal omasal, la proportion d’eau réellement absorbée dans le feuillet est de 8 %. Environ 50 % des AGV qui pénètrent dans le feuillet sont absorbés. La concentration des micro-organismes en µg ADN /ml est de 60 % supérieure à celle du rumen avec une activité fermentaire moindre et qui a une vitesse de 50 % inférieure à celle du rumen (Giesecke et Engelhardt 1975 cités par Deswysen et al., 1995) .Par contre la concentration des protozoaires n’est que de : • 14 % du flux d’entrée omasal chez le mouton (Weller et Pilgrim 1974 cités par Deswysen et al., 1995) • 11 à 17 % du contenu omasal interlamellaire (Town et Nagaraja 1990 cités par Deswysen et al., 1995). • 6.7 % de l’effluent omasal chez le mouton (Mc Sweeny, 1986 ) Tableau n° 8 : Absorption et sécrétion de différents électrolytes et minéraux dans le feuillet [de 04 moutons et 02 chèvres (Von Engelhardt et Hauffe, 1975 b) et de 04 jeunes bovins ( Edrise et Smith , 1979) en % de l’effluent réticulaire pénétrant dans le feuillet] cités par Deswysen et al., 1995. 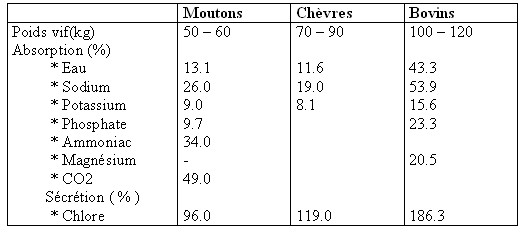 III.2- Dans la caillette : Chez les ruminants , les aliments solides sont profondément dégradés dans le rumen-réseau sous l’action des micro-organismes et les corps bactériens constituent une part très importante de produits qui arrivent dans la caillette .Les digesta arrivent librement dans la caillette et y séjournent très peu (30 à 60 minutes ).Du fait du transit plus régulier et de la sécrétion plus importante d’acide chlorhydrique , le pH de la caillette fluctue moins et est en moyenne plus bas ( 2 à 3 ).Ces conditions sont plus favorables à la dénaturation des matières azotées et à l’hydrolyse des protéines sous l’action de la pepsine mais le temps de séjour très bref a l’effet inverse .L’acidité du milieu tue les protozoaires .Le lysozyme joue un rôle important chez le ruminant puisqu’il est sécrété en abondance , il intervient dans la lyse des parois bactériennes et facilite ainsi la dissociation des bactéries fixées et des particules .On estime chez le mouton entre 30 à 70 % des microbes ( bactéries et protozoaires ) fixés aux particules provenant du rumen qui sont détachées dans la caillette (Yang, 1991 cité par Toullec et Lalles, 1995). L’absorption porterait aussi sur les AGV non absorbés dans le feuillet (ITEB –INRAP, 1984). III.3- Dans l’intestin grêleLes mécanismes de digestion et de l’absorption dans l’intestin grêle sont les mêmes que chez les monogastriques .Les enzymes du suc pancréatique (les trypsines ,les chymotrypsines A ,B , C , l’élastase , les carboxypeptidases A , B , la lipase plus colipase , la cholestérol estérase , l’amylase , la ribonucléase , la désoxyribonucléase ) et de la muqueuse intestinale( aminopeptidase A et N , la phosphatase alcaline , la lactase , la maltase et l’isomaltase ) sont les mêmes , par contre les substances pénétrant dans l’intestin grêle sont différentes .Il faut toutefois noter que le pancréas des ruminants secrète beaucoup plus de nucléases que celui des mono-gastriques III.3.1- Les glucides : on trouve à l’entrée de l’intestin grêle :*de l’amidon qui est hydrolysé en maltose puis en glucose et il est absorbé .La quantité de glucose absorbée représente moins de 1 % de l’énergie des nutriments absorbés avec les fourrages seuls. Elle ne devient notable qu’avec des rations mixtes contenant du mais et du sorgho. Le ruminant doit donc synthétiser la quasi-totalité ou la majeure partie du glucose dont il a besoin. On estime que l’apport alimentaire de glucose fournit en moyenne 5 % de l’énergie absorbée. *des glucides pariétaux non dégradés dans le rumen – réseau et qui ne seront pas aussi dégradés au niveau de l’intestin grêle . *Les résidus des polyosides de réserve des bactéries ( ITEB –INRAP, 1984). III.3.2- Les protéines Les protéines qui arrivent au niveau du duodénum sont de deux types :alimentaires et microbiennes. Il est admis que 80 % des protéines microbiennes se trouvent sous forme de protéines vraies et 20 % sous forme d’acides nucléiques qui n’auraient aucune valeur pour l’animal (ITEB-INRAP, 1984). La digestion et l’absorption des protéines auraient lieu principalement dans le deuxième tiers de l’intestin grêle ; elles seraient donc faibles dans l’iléon malgré ses capacités élevées entre autre un pH de 7.5 favorable à l’action des enzymes protéolytiques (Toullec et al., .,1995). L’analyse de 405 bilans réalisés sur des moutons ,jeunes bovins et vaches laitières recevant des rations mixtes ou des fourrages verts distribués seuls , le flux d’azote à l’entrée du duodénum est pour • 56 % d’origine microbienne (37 à 88 % ) • 38 % d’origine alimentaire (10 à 60 % ) • * 6 % d’origine endogène (Toullec et al., 1995). La digestibilité des protéines dans l’intestin grêle varie considérablement de 0.5 à 0.80 . (Vérité et al., 1987 ). Elle est estimée à 0,80 pour les protéines d’origine microbienne et de 0.25 à 0.95 pour les protéines alimentaires. Pour un niveau d’alimentation donné , elle est liée positivement au flux duodénal de protéines et négativement au flux iléal de matière non protidique (Van Bruchen et al., 1985 cités par Toullec et al., 1995). Les protides qui échappent à la digestion dans l’intestin grêle sont donc constitués principalement de fractions communes d’origines bactérienne et endogène , auxquelles vient se rajouter une fraction d’origine alimentaire ,moins importante et de composition variable (Greife et al., 1985 et Lallès et al., 1990 tous deux cités par Toullec et al., 1995). III.3.3- Les lipides :Les lipides sont efficacement digérés dans l’intestin , grâce à la bile et au suc pancréatique ; la bile joue un rôle particulièrement important en apportant des phospholipides qui facilitent la mise en solution micellaire des acides gras insaturés .Dans le duodénum ,l’apport de lipides par la bile entraîne une augmentation marquée des quantités d’acides gras insaturés ,de phospholipides et de cholestérol (Noble, 1981 ; Bauchart, 1993 ; Doreau et al., 1994 tous cités par Toullec et al., 1995). Chez le mouton , la quantité totale de lipides ainsi sécrétée équivaut à environ 50 % de la quantité ingérée avec des rations classiques non supplémentées en lipides . La digestion des lipides se produit dans un milieu bi-phasique constitué d’une phase particulaire insoluble et d’une phase micellaire soluble. Cependant et contrairement à ce qui est observé chez les pré-ruminants ,la phase insoluble n’est pas une émulsion de globules gras stabilisée par les phospholipides et les sels biliaires , puisque les lipides sortant de la caillette sont liés à des débris végétaux , à des corps microbiens ou à des cellules desquamées. Les acides gras non estérifiés sont progressivement transférés dans la phase micellaire soluble , grâce à l’action détergente des lyso-lécithines et des sels biliaires. Cependant très peu de données sont disponibles sur la digestibilité des lipides dans l’intestin grêle (Doreau et al., 1994 , Toullec et al., 1995 ). Le plus souvent , les mesures ont été effectuées à l’entrée du duodénum et dans les fèces or le passage dans le gros intestin entraîne des modifications profondes et variables . | |
| | | | amira
Messages : 72
Points : 26
| | Sujet: Re: COURS DE NUTRITION ANIMALE DEUXIEME ANNEE (Par DR MEZIANE TOUFIK) Ven 21 Aoû 2009 - 11:14 | |
| III.4 – Dans le gros intestin Le contenu du gros intestin constitue un milieu très favorable pour le développement d’une digestion microbienne .La température y est constante et le pH , peu variable ,reste voisin de la neutralité (6.6 à 7.8 ).Le cæcum se remplit par intermittence chez le mouton : le flux digestif sortant de l’iléon terminal peut atteindre de 70 à 90 g en 10 mn , être suivi d’un arrêt de 30 à 60 mn , et n’est pas en liaison directe avec les activités d’ingestion ou de rumination (Ulyatt et al., 1975 cités par Tisserand et Demarquillly 1995 ). Des contractions coordonnées péristaltiques et antipéristaltiques assurent le mélange du contenu du cæcum à la cadence de 3 à 7 par période de 10 mn (Mac Rue et al., 1973 cité par Tisserand et al., 1995) .Des vagues de 2 à 5 contractions intenses à des intervalles de 30mn à 4 heures assurent la vidange du cæcum (Ruckebush 1970 cité par Tisserand et al., 1995) Les céréales diminuent l’activité du cæcum chez les moutons et les bovins et ce vraisemblablement par suite d’une fermentation cæcale accrue ou anormale .Le temps de séjour des digesta dans le gros intestin du mouton varie de 10 à 29 heures , comparable à celui de la phase liquide du rumen réseau, mais plus court que celui de la phase solide dans le même compartiment. Il diminue quant le niveau de l’ingestion augmente .( Ruckebush 1970 et Svendsen 1972 cités par Tisserand et al.,(1995). La population microbienne du gros intestin est caractérisée par des bactéries peu différentes de celles du rumen - réseau mais en quantités moins importantes (Ulyatt et al., 1975 , Hoover 1978 cités par Preston et al.,1987 et par Tisserand et al.,1995). La présence de protozoaires reste incertaine et il existe peu de données sur les champignons bien que certains auteurs signalent leur présence dans le gros intestin des grands ruminants (Kern et al.,1974 ,Allison et al.,1975 cités par Tisserand et al., 1995) Tableau n° 9: Temps de séjour des aliments dans le gros intestin ( Tisserand et al., 1995) 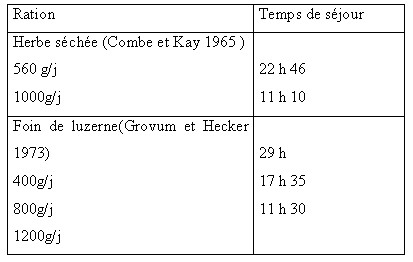 III.4.1- Les glucides III.4.1- Les glucides Seuls les glucides pariétaux (cellulose et hémicelluloses) non digérés dans le rumen arrivent dans le gros intestin , aucun glucide soluble n’est détecté dans les digesta de l’iléon lorsqu’on est en présence d’une ration à base de fourrages classiques . (Beever et al., 1972 cités par Tisserand et al., 1995). Cependant , la quantité de glucides solubles arrivant dans le gros intestin peut augmenter avec des rations à base de céréales et surtout le mais et le sorgho où une grande quantité d’amidon peut échapper à la dégradation ruminale .La population microbienne du cæcum et du colon utilise ces glucides en produisant des acides gras volatils comme dans le rumen mais avec une quantité moindre .Les AGV absorbés à ce niveau représentent une fraction très variable , de 4 à 26 % de l’énergie totale digérée (Thomas et Rook 1977 cités par Tisserand et al., 1995). Tableau n°10 : Pourcentage molaire des AGV dans le côlon et le cæcum (Faicheney 1969 cité par Tisserand et al., 1995). 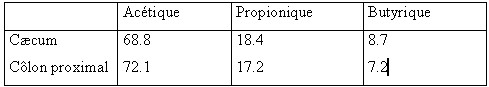 D’après Beever et al., 1972 cités par Tisserand et al., 1995, la digestibilité de la cellulose et des hémicelluloses dans le gros intestin de mouton (en % des quantités qui y entrent ) est estimée de : • 18.5 à 49.5 % pour la cellulose • 24.9 à 48.1 % pour les hémicelluloses Cependant les quantités qui y sont digérées , représentent de : • 3.0 à 29.6 % pour la cellulose • 7.9 à 41.1 % pour les hémicelluloses , des quantités digestibles dans la totalité du tube digestif. III.4.2-Les protéinesLa quantité d’azote qui arrive dans le cæcum représente : 25 à plus de 50 % de la quantité d’azote ingérée (surtout avec les rations pauvres en azote ) Ulyatt et al., 1975 cités par Tisserand et al., (1995). La moitié de cet azote est sous forme soluble, jusqu’à 17 % sous forme ammoniacale .Les protéines représentant 40 à 80 % de cet azote ,sont d’origine alimentaire , microbienne et endogène auxquelles il faut ajouter l’azote provenant du mucus sécrété par la paroi du gros intestin , les cellules desquamées et surtout l’urée provenant du sang et diffusant à travers la paroi. (Tisserand et al., 1995) Les bactéries du cæcum et du côlon proximal ont une importante activité de protéolyse , de désamination , de décarboxylation et d’uréolyse. La dégradation des protéines aboutit à la formation des acides aminés eux mêmes dégradés en ammoniac , acides gras volatils et méthane (Mason et al., 1981). Selon Dixon et Nolan (1982) cités par Tisserand et al.,( 1995) , plus de 3 g de azote ammoniacal sont produits par jour dans le cæcum et le côlon proximal du mouton dont un quart provient de l’urée endogène. D’après Mac Neil 1988 cité par Tisserand et al., (1995) , le métabolisme de l’azote dans le gros intestin apporte peu d’acides aminés à l’animal ( 6 % maximum des apports ) mais le gros intestin participe de façon importante au reecyclage de l’urée participant de ce fait pour 40 % au transfert net d’azote de l’ensemble du tube digestif aux liquides corporels. Tableau n° 11 :Transfert net d’azote du tube digestif aux liquides corporels chez le mouton recevant des fourrages (Mazanov et Nolan 1976 cités par Tisserand et al., 1995)  Le gros intestin qui n’a fait l’objet que d’un nombre réduit d’études, comparativement au rumen, joue apparemment un rôle important dans le recyclage de l’ammoniaque avec un rôle non négligeable dans la digestion des parois végétales dans le cas d’une diminution du temps de séjour des digesta dans le rumen suite à un niveau alimentaire élevé ou aussi lors de la diminution de l’activité cellulolytique entraînée par des régimes riches en amidon rapidement fermentescible.( Tisserand et al., 1995 ) III.4.3-Absorption d’eau et de minéraux . Le gros intestin joue un rôle important dans l’absorption de l’eau et de certains minéraux majeurs comme chez toutes les espèces . Chez les ruminants environ 90 % de l’eau entrant dans le gros intestin du mouton y est absorbée, la teneur du contenu du gros intestin décroît régulièrement de 7.1g d’eau par g de MS dans le cæcum à 1.5g dans le rectum chez le mouton [Kay et Pfeffer., 1970 , Grovum et Hecker., 1973] cités par Tisserand et al., (1995) . Le gros intestin est aussi un lieu d’absorption de nombreux éléments minéraux avec une prédominance d’absorption du sodium ,en grande partie d’origine endogène ,du phosphore, calcium ,et magnésium ainsi que le chlore alors que le potassium n’y est que faiblement absorbé .Il existe aussi une absorption importante des oligo-éléments comme le zinc ,cobalt, cuivre et du manganèse (Hoover 1978 cité par Tisserand et al., (1995) | |
| | | | amira
Messages : 72
Points : 26
| | Sujet: Re: COURS DE NUTRITION ANIMALE DEUXIEME ANNEE (Par DR MEZIANE TOUFIK) Ven 21 Aoû 2009 - 11:30 | |
| IV- UTILISATION METABOLIQUE DES NUTRIMENTS A la fin de la digestion , le contenu digestif se réduit aux substances provenant des aliments et des corps microbiens .Les nutriments résultant de la digestion sont absorbés et transportés aux cellules où ils seront utilisés. IV.1-L’absorption Elle s’opère le long des muqueuses du tractus digestif et principalement : a - La muqueuse du rumen-réseau absorbe les AGV provenant de la dégradation de la cellulose , l’ammoniac en excès , mais avec des risques d’intoxication en cas de forte absorption ,en plus de l’absorption de certains minéraux (Mg ,S , P ,Ca )et la sécrétion d’autres éléments minéraux (P ,Ca ,Na) (Meschy et al., 1995, Rémond et al., 1996). b- La muqueuse du feuillet absorbe beaucoup d’eau et de sels minéraux (Mg , Na) Leur résorption favorise l’acidification des aliments dans la caillette . Les muqueuses du feuillet et de la caillette absorbent le reste des AGV qui n’auraient pas eu le temps d’être absorbés par le rumen , soit 18 à 30 % du total.(Soltner 1994). c- La muqueuse de l’intestin grêle absorbe tous les nutriments (glucose ,glycérol , acides gras à longues chaînes issus des corps gras ,acides aminés , minéraux (Ca ,P ,K, Cl), vitamines et eau . Son rôle est surtout important chez les mono-gastriques . d- La muqueuse du gros intestin absorbe surtout de l’eau et des sels minéraux (NaCl, Mg, Ca) , des AGV (essentiellement de l’acide acétique ), de l’acide lactique et du NH3. L’énergie absorbée représente moins de 10% de celle absorbée par le rumen et l’intestin grêle. Par contre la totalité des corps microbiens formés dans le gros intestin sont excrétés avec les déjections dont il représente la majeure partie de l’azote (Soltner 1994). IV.2-L’utilisation métabolique Les nutriments énergétiques et plastiques sont transportés par le sang jusqu’au foie , puis apportés aux cellules où ils vont participer à une multitude de réactions chimiques nécessaires à la vie et constituent le métabolisme. Ce dernier revêt deux aspects liés entre eux : l’anabolisme et le catabolisme . Les nutriments énergétiques : les ruminants tirent leur énergie des acides gras volatils (AGV) issus de la dégradation de la cellulose ,ils constituent la principale source d’énergie des ruminants 66 à 75 % de l’énergie disponible pour l’organisme.L’énergie des AGV , du lactate et des corps cétoniques (ß -OH-butyrate et acéto-acétate ) représentent environ 50 % de l’énergie métabolisable ingérée. Ces valeurs sont inférieures chez les ovins (brebis taries ,gestantes ou en lactation) [ Lindsay, 1993 cité par Journet 1995]. A cet effet , le taux de glucose sanguin des ruminants est inférieur à celui des mono-gastriques alors que leur taux d’acides gras volatils est plus élevé .Il faut noter que les AGV absorbés sont partiellement métabolisés par l’épithélium ruminal . Environ 30 % de l’acétate , 50 % du propionate ,et 75 à 85 % du butyrate produits dans le rumen sont utilisés ou métabolisés par la paroi du tube digestif [Bergman et Wolfe 1971 ,Weekes et Webster 1975 ]cités par Journet et al., 1995. Les principaux produits formés retrouvés dans la circulation sanguine sont : le lactate issu du propionate et du ß -OH-butyrate issu du butyrate . Krehbiel et al., 1992 cités par Journet et al., 1995, estiment que seulement 30 % du butyrate produit entrent dans le système porte ; alors que Seal et Parker, 1994 cités par Journet et al., 1995 confirment que plus de 50 % du propionate sont métabolisés dans le rumen. IV.2.1-Le métabolisme des substances non azotées : Il est basé sur l’utilisation du glucose , des acides gras volatils ,et des acides gras qui fournissent la majeure partie de l’énergie mise à la disposition de l’organisme .Leur métabolisme correspond en grande partie au métabolisme énergétique du ruminant. Toutefois, une part variable de l’énergie absorbée provient de l’utilisation énergétique des nutriments azotés ,les acides aminés .Les nutriments utilisés à des fins énergétiques ont deux fonctions . *Par le catabolisme oxydatif , ils fournissent de l’énergie sous forme d’ATP utilisable par les systèmes enzymatiques cellulaires comme fournisseurs d’énergie nécessaire aux biosynthèses (anabolisme ).Ces nutriments empruntent les voies métaboliques telles que la glycolyse, le cycle de Krebs ou l’oxydation des acides gras à longues chaînes. *Par anabolisme énergétique , ils permettent la biosynthèse des constituants du lait ou des tissus. Les voies métaboliques sont en relation les unes avec les autres, la voie métabolique du glucose en relation avec celle des triglycérides (lipides) et de certains acides aminés pouvant être utilisés à des fins énergétiques .Ces interrelations se font au niveau de certains carrefours importants permettant la connexion des voies métaboliques , trioses phosphates et puryvate . Tableau n° 11: Utilisation des différents substrats par les principaux tissus de l’organisme (Remesy et Demigne 1981, ITEB –INRAP 1984) 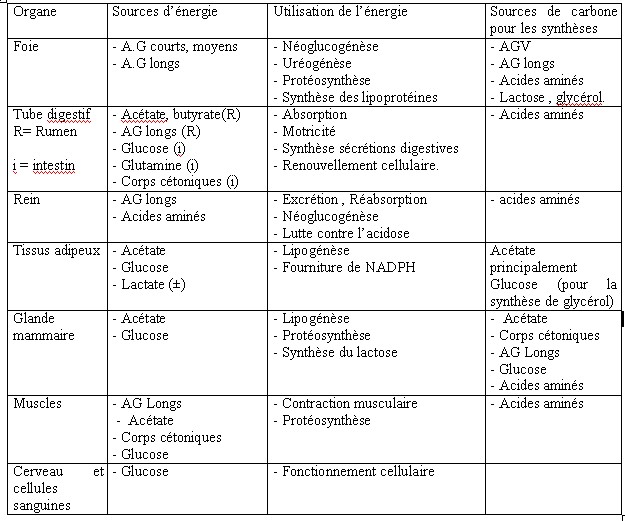 IV.2.1.1- Le métabolisme glucidique IV.2.1.1- Le métabolisme glucidique Ce métabolisme suit les voies métaboliques : a-La glycolyse conduit à l’acide pyruvique avec un carrefour important constitué par les trioses phosphates qui sont les précurseurs du glycérol lui même à la base de la synthèse des lipides corporels . b-Le Cycle de Krebs ou cycle citrique , ou cycle tri-carboxylique : il constitue le système qui permet de dégrader les produits terminaux des métabolismes des oses , des acides gras et de nombreux acides aminés en permettant la production de al plus grande partie de l’énergie dont les cellules ont besoin. Comme la glycolyse, il fournit de l’énergie .De par son rôle , le glucose fournit : -l’énergie aux cellules nerveuses , aux cellules des muscles lisses et des globules rouges , une grande partie ( 30 à 40 % )de l’énergie nécessaire au fonctionnement de la mamelle. -Il est le précurseur obligatoire du lactose , -Il peut aussi servir à la synthèse du glycérol et des acides gras , donc des triglycérides corporels , -Il est le substrat énergétique essentiel et le principal précurseur des lipides chez le fœtus .( ITEB -INRAP, 1984). Or le glucose ne constitue pas le nutriment énergétique le plus important chez les ruminants , il ne représente en effet que 5 % en moyenne de l’énergie absorbée (moins de 1 % pour les fourrages seuls ) (Vermorel., 1981) . L’origine du glucose entrant dans le métabolisme est double chez le ruminant : • l’absorption intestinale du glucose est limitée à 15 % du total. • la fourniture des 85 % restants est assurée par principalement dans le foie par une synthèse appelée la néo-glucogénèse hépatique et à un moindre degré la néo-glucogénèse ( la NGG) rénale , à partir de substances gluco-formatrices (Vermorel., 1981; Rémésy et Demigne, 1981 ; Payne 1983 ; ITEB- INRAP, 1984 ; Preston et al., 1987). Il semble d’après Huntington et Reynolds, 1986 cités par Journet et al.,1995, qu’une proportion faible (de 30 à 35 % ), du glucose issu de la digestion intestinale de l’amidon se retrouve dans le sang porte à la suite d’infusions d’amidon dans le duodénum alors que cette proportion atteint 64 à 94 % pour le glucose infusé à ce niveau Tableau n° 12 : Utilisation des principaux substrats gluco-formateurs au niveau du foie . (INRA 1978 ) 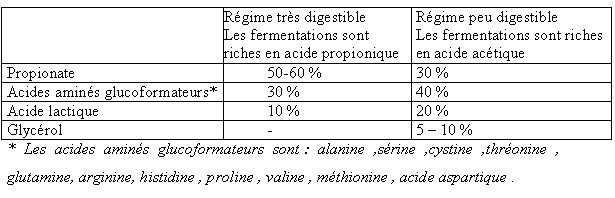 Ces essais montrent que le tube digestif dont le métabolisme est très intense , prélève une quantité importante de glucose. Le métabolisme énergétique concerne un très grand nombre de métabolites ,dont beaucoup sont utilisés par les cellules comme combustible pour la respiration oxydative . Le glucose est considéré d’ordinaire comme le combustible le plus important ,certainement vrai pour les non ruminants et même pour les ruminants dont il reste indispensable pour certaines fonctions clés comme le métabolisme cérébral et la lactation . Le métabolisme énergétique des ruminants est constitué du métabolisme glucidique et lipidique . Chez les ruminants , le glucose circulant résulte de l’absorption intestinale de glucose et de la néo-glucogénèse (NGG). Ce glucose provient aussi de l’amidon qui a échappé à la dégradation ruminale et aussi de glucosanes de réserves des micro-organismes du rumen-réseau. La glycémie est faible au cours d’un jeûne ( à partir du deuxième jour de jeûne ) (Ndibualonji et al., 1997 ) si l’apport est insuffisant en énergie (par manque d’aliments précurseurs du glucose ) ou aussi lors de carence en cobalt (car chez les ruminants la vitamine B12 conditionne l’utilisation de l’acide propionique pour la néo-glucogénèse) (Haddad, 1981; Jouany et al., 1995). Les états d’hyperglycémie chez les ruminants peuvent être rencontrés chez les animaux recevant des rations à base d’ensilage de mais que chez ceux alimentés au foin ou lors de consommation d’aliments concentrés(Rowlands et al., 1974 ). Selon Haddad, 1981, le niveau d’ingestion n’a d’effet ni sur la glycémie moyenne journalière ni sur l’évolution durant la journée. Tableau n° 13 : Normes physiologiques de la glycémie chez le mouton 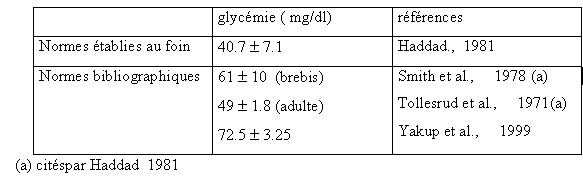 | |
| | | | amira
Messages : 72
Points : 26
| | Sujet: Re: COURS DE NUTRITION ANIMALE DEUXIEME ANNEE (Par DR MEZIANE TOUFIK) Ven 21 Aoû 2009 - 11:44 | |
| Facteurs de variations Des variations peuvent exister : - entre les troupeaux de bovins. - L’âge : la glycémie diminue chez l’agneau dés la deuxième semaine et se stabilise au niveau de l’adulte aux environs de quatre mois. - La saison : le taux augmente en mai et en novembre. - Chez les femelles ruminants dans les heures qui suivent le vêlage, la glycémie peut atteindre 2 g/l sous l’action des catécholamines (Sauvant,1980 cité par Haddad, 1981). - La concentration du glucose sanguin est diminue en fin de gestation , à la suite de l’utilisation du glucose par l’utérus (Rémond et al., 1973 ; Grizard, 1979) cités par Haddad, 1981). - La concentration du glucose sanguin est minimale au début de la lactation ,et augmente ensuite chez les vaches laitières. - Chez la brebis , il a été noté un accroissement rapide de la glycémie juste après le part.(Scott et al., 1976 cités par Haddad. 1981) - L’hypoglycémie de nutrition a pour effet de provoquer des anoestrus et d’augmenter l’incidence d’acétonémie et d’infertilité (Payne et al., 1974 , Savaria 1975 cité par Haddad 1981). - Selon plusieurs auteurs cités par Payne, 1983, en cas de sous alimentation grave, les acides gras libres plasmatiques peuvent être multipliés par cinq. Alors que, le taux de glucose passe de la valeur normale 45 mg/100ml à une valeur bien souvent inférieure à 40 mg /100ml. Ces résultats doivent être interprétés cependant avec prudence dans les troupeaux soumis à des carences énergétiques marginales . VI.1.1.2-Le métabolisme des AGVLes AGV absorbés au niveau de la paroi du rumen constituent la principale source d’énergie pour le ruminant, 60 à 80 % de l’énergie mise à la disposition de l’organisme( ITEB –INRAP, 1984). L’absorption des tous les AGV est facilitée par les papilles de la muqueuse du rumen qui augmentent considérablement sa surface et ses capacités d’absorption. Les aliments grossiers qui produisent beaucoup d’acide acétique dans le rumen ont tendance à stimuler le développement de ces papilles et augmentent ainsi la capacité d’absorption. L’absorption des AGV ne se fait pas activement mais selon un processus passif en fonction d’un gradient de concentration, le taux d’absorption est proportionnel à la concentration à l’intérieur du rumen. Le pH intervient aussi et le taux d’absorption est particulièrement rapide lorsque le jus du rumen est acide (Payne, 1983 ; ITEB –INRAP, 1984 ). Dans les conditions normales d’alimentation, la composition des mélanges d’acides organiques présents ou produits dans le rumen et de ceux qui sont absorbés (absorption nette dans le sang porte ) n’a fait l’objet que de très peu d’études . La proportion d’acétate s’accroît aux dépens de celle du propionate et surtout du butyrate qui sont métabolisés par l’épithélium ruminal en donnant naissance à de nouveaux produits tels l’acide lactique et l’acide ß -OH-butyrique. Les proportions des acides organiques absorbés passent pour : • *L’acide acétique de 60 à 70 % avec les rations à base d’ensilages de luzerne ou de dactyle (R1)à 45 % avec des rations contenant 85 % d’aliments concentrés (R2) • *L’acide propionique : de 16 à 24 % (R1) à 25 - 35 % (R2) • *L’acide L-lactique : 6-9 % ( R1) à 12 –15 % (R2) l’acide lactique peut aussi provenir de l’oxydation incomplète du glucose (Payne, 1983 ). Remarque : la proportion du propionate et du lactate augmente avec les rations contenant de l’amidon fermentescible(Gross et al., 1988, ITEB-INRAP 1984 ). a -Le propionate pénètre le cycle de Krebs au niveau du succinate et il y est oxydé pour fournir de l’ATP. Son utilisation métabolique est très intéressante, il peut emprunter des voies métaboliques variées : Il peut permettre la formation de glucose par une des voies de la néoglucogénèse où il est utilisé préférentiellement au niveau du foie pour fournir le glucose nécessaire au ruminant et que celui-ci ne peut trouver qu’en quantité généralement très faible à partir de l’hydrolyse des glucides des aliments Il est donc glucoformateur et constitue la principale source de glucose endogène pour les ruminants consommant des rations très digestibles. La néoglucogénèse (NGG) est donc un phénomène capital chez le ruminant dont le besoin (en g /kg de poids métabolique / h ) est comparable à celui déterminé chez le monogastrique .L’activité des enzymes de la NGG (pyruvate carboxylase, phosphoénolpyruvate carboxykinase) est d’ailleurs différente et considérablement plus importante dans la mitochondrie chez le ruminant que chez le monogastrique (Ballard et al., 1969 cités par Thivend et al., 1985 ).En alimentant le cycle de Krebs en oxaloacétate , le propionate joue un rôle anticétogène . Le propionate permet aussi le formation de glycérol , par le carrefour des trioses phosphates , alimentant la synthèse des triglycérides . Il permet aussi la formation d’acétyl-CoA, par le carrefour du pyruvate ; l’acétyl CoA pouvant lui même servir à la synthèse des acides gras longs. Tableau n° 14 : Origine et répartition (en %) de l’énergie absorbée dans le tube digestif du ruminant (Vermorel 1978 cité par Thivend 1985)  b- Le butyrate est métabolisé en grande partie au niveau de la paroi du rumen en corps cétoniques(et en ß -OH-butyrate en particulier), il est donc cétogène . Les corps cétoniques ont un taux dans le sang de la vache entre 5 et 10 mg/100ml .Le butyrate restant est métabolisé dans le foie où il est glucoformateur . c-L’acide acétique : Outre son utilisation directe par la mamelle pour la synthèse des acides gras courts et moyens des matières grasses du lait , l’acide acétique est métabolisé rapidement en acétylcoenzyme A ou acétyl Coa qui est une molécule organique riche en énergie , qui occupe une position clé dans les métabolismes , comme introducteur d’acides gras . Les destinées de l’acétate se lient à l’acétyl selon les voies métaboliques suivantes : La voie 1 : l’acétate peut entrer dans le cycle de Krebs où il est oxydé et fournit de l’énergie (ATP) , il faut pour cela qu’il y ait suffisamment d’acide oxalo-acétique dans le cycle pour alimenter la combustion La voie 2 : il peut servir de point de départ à la biosynthèse des acides gras à longue chaîne et donc alimenter un des éléments de la synthèse des lipides ou lipogénèse La voie 3 : cette voie est empruntée quand les voies 1 et 2 sont saturées ou impossibles. L’acide acétique donne naissance alors , via l’acétyl CoA , à des corps cétoniques qui peuvent être utilisés à des fins énergétiques s’ils ne sont pas en excès ; s’ils le sont ils deviennent alors toxiques et provoquent l’acétonémie ou acétose . On dit que l’acide acétique est cétogène. L’acétyl CoA peut aussi servir de précurseurs à la synthèse du cholestérol , lui même précurseur des acides biliaires , des hormones stéroïdes et de la vitamine D3. La voie 4 : représente la réaction pyruvate ________acétyl CoA qui est irréversible .L’acide acétique ne peut donc pas servir à la synthèse du glucose par la néoglucogénèse ; il n’est pas glucoformateur. Taux d’acétate dans le sang chez les bovins : 10 mg /100ml 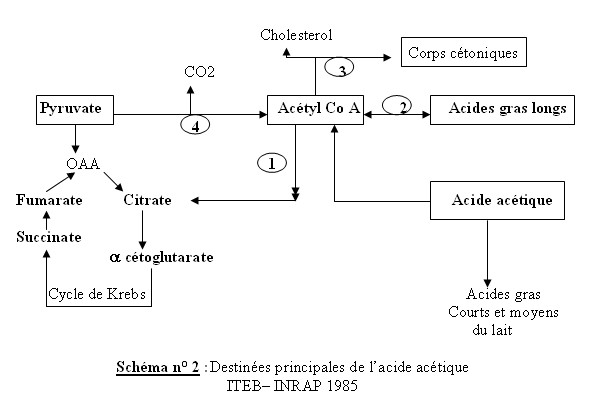 | |
| | | | amira
Messages : 72
Points : 26
| | Sujet: Re: COURS DE NUTRITION ANIMALE DEUXIEME ANNEE (Par DR MEZIANE TOUFIK) Ven 21 Aoû 2009 - 12:09 | |
| VI.2.1.3- Le métabolisme des lipides Au niveau des tissus , il existe simultanément deux phénomènes antagonistes , la lipogenèse ou synthèse des lipides corporels , et la lipolyse ou destruction des lipides corporels . Les acides gras des lipides sont absorbés au niveau de l’intestin grêle sous forme de micelles contenant des lyso-lécithines biliaires ; une partie de l’acide stéarique est désaturée en acide oléique dans l’épithélium intestinal (Bickerstaffe et al., 1972 cités par INRA 1978 , Remesy et al., 1984 ).Les acides gras absorbés sont estérifiés en triglycérides , transportés par la lymphe et déversés dans le sang sous forme de chylomicrons utilisés par les tissus adipeux et la glande mammaire (Aurosseau. 1981). VI.2.1.3.1– Les lipides totaux Une grande partie des lipides est représentée par les triglycérides , le cholestérol , les phospho-lipides et les acides gras éstérifiés .Ces fractions sont combinés aux protides pour former les lipoprotéines .L’autre partie se compose des acides gras non éstérifiés qui proviennent de l’hydrolyse du tissu adipeux . Le taux des lipides du sang dépend essentiellement de la composition des aliments, il y aurait une augmentation de la teneur des lipides sanguins après l’absorption d’un repas riche en graisses.(Haddad 1981) Tableau n° 15 :Normes physiologiques de la lipémie chez le mouton  Facteurs de variations Facteurs de variations La lipémie dépend : - De l’âge et du sexe : selon Vermorel (1981) , les femelles sont plus précoces que les males castrés et plus encore que les males entiers dans le développement du tissu adipeux . - Les productions ( la gestation et la lactation ) semblent avoir une influence importante . Chez les vaches laitières , on observe une teneur plus élevée en lipides que chez les autres types de vaches (allaitantes ou taries). La quantité des acides gras prélevés dans le courant sanguin peut s’accroître quand les les lipides circulants augmentent en particulier lors de mobilisation des réserves énergétiques en début de lactation .Cette augmentation porte sur les acides gras non estérifiés [(Enjalbert, 1994 ; Raphael et al., 1973) cités par Haddad, 1981 ; Sauvant et al., 1978 ; Haddad, 1981]. - Les états d’inanition chronique (cachexie ) provoquent une diminution de la lipémie (Haddad, 1981). VI.2.1.3.2- Les triglycéridesDans les cellules intestinales, la majorité des acides gras sont unis au glycérol (provenant du sang) pour former des triglycérides. Ces triglycérides, certains acides gras libres, le cholestérol et d'autre substances lipidiques sont couvertes d'une protéine pour former les lipoprotéines riches en triglycérides (LP-TG), aussi appelées chylomicrons ou lipoprotéines de très faible densité. Les lipoprotéines riches en triglycérides sont absorbées dans les vaisseaux lymphatiques. Ils rejoignent la circulation générale au niveau de la jonction thoracique (la jonction entre le système lymphatique et le système sanguin). Contrairement à la plupart des autres nutriments, les lipides entrent dans la circulation sanguine générale et sont utilisés par les tissus du corps sans être d'abord métabolisés par le foie. En cas de sous alimentation sévère , un des phénomènes essentiels est constitué par la dégénérescence graisseuse du foie , le contenu lipidique total du foie double , le taux des triglycérides augmente vingt fois, celui du cholestérol estérifié huit fois et celui des acides gras libres trois fois. Le foie est incapable d’augmenter la sécrétion de lipoprotéines capables de transporter cette graisse en dehors du foie (lipotropie). La surcharge hépatique est un facteur important à considérer pendant la sous alimentation, mais aussi pendant le rétablissement et la réalimentation (Payne, 1983). VI.2.1.3.3- Le cholestérol Dans le sang , le cholestérol est toujours lié à une protéine et à une ou plusieurs molécules de phospho-lipides formant la lipoprotéine .Il se présente sous deux formes estérifiée (70 %) et non estérifiée (30 %) . Il a une double origine ; alimentaire et endogène .Il est surtout synthétisé dans le foie et également dans l’intestin , les surrénales , les testicules , les ovaires , la peau et le système nerveux (Haddad. 1981 , Sommer H. 1984) . Tableau n° 15 : Normes physiologiques de la cholestérolémie chez le mouto 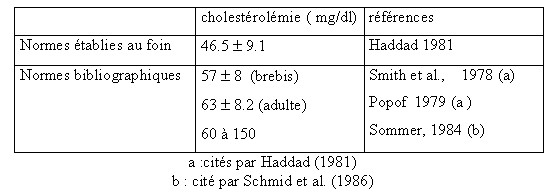 Facteurs de variations Facteurs de variations La cholestérolémie est généralement stable mais peut être influencée par : - Une ration alimentaire à base de concentrés entraîne l’augmentation de la cholestérolémie qui peut atteindre 2.39 mmol/l . Cette augmentation semble suivre la même évolution que la lipémie . - La saison, la région, l’âge. En pathologie , on peut noter : *L’hypercholestérolémie lors de syndrome néphrotique, hypothyroidisme , les maladies du foie (cirrhose ), lors de corticostéroidothérapie ou lors d’hyperlipidémie (Sommer, 1984 ) ou lors d’ictère par retention (Haddad, 1981). L’hypocholestérolémie est rencontrée lors du tarissement et en période puerpérale et lors de cachexie ,d’hyperthyroidisme. Schéma n° 3 : Schéma simplifié du métabolisme énergétique des produits terminaux de la digestion dans l’épithélium du tube digestif et le foie (INRA, 1978 )  à suivre | |
| | | | amira
Messages : 72
Points : 26
| | Sujet: Re: COURS DE NUTRITION ANIMALE DEUXIEME ANNEE (Par DR MEZIANE TOUFIK) Ven 28 Aoû 2009 - 17:41 | |
| VI.2.2- Le métabolisme des substances azotées.
VI.2.2.1- Les protéines totales et albumine Les protéines fournissent les acides aminés nécessaires pour le maintien des fonctions vitales, la croissance, la reproduction et la lactation. Les animaux non-ruminants ont besoin d'acides aminés préformés dans leur ration. Par contre, grâce aux microbes présents dans le rumen, les ruminants possèdent la capacité de synthétiser les acides aminés à partir d'azote non-protéique (ANP). Des sources d'ANP telles que l'ammoniac ou l'urée peuvent donc être utilisées dans leur ration. De plus, les ruminants possèdent un mécanisme pour conserver l'azote lorsque leur ration est déficiente en azote. L'urée est le produit final du métabolisme des protéines dans le corps et elle est normalement sécrétée dans les urines. Cependant, en cas de déficit azoté, l'urée retourne de préférence dans le rumen où les bactéries peuvent en faire usage (Payne, 1983 ; Ndibualonji et al. 1997). Chez les non-ruminants, l'urée produite dans le corps est toujours entièrement perdue dans les urines (Wattiaux 1996). Les protéines plasmatiques forment en fait un mélange très complexe comprenant non seulement des protéines simples, mais aussi des formes conjuguées telles que les glycoprotéines et différents types de lipoprotéines .L’emploi de différentes concentrations de sulfates de sodium ou d’ammonium permet habituellement de séparer les protéines de plasma en trois groupes principaux : fibrinogène, albumine et globulines. La proportion des diverses fractions des protides sériques varie selon l’espèce animale , tandis que chez l’homme , le taux des albumines est plus élevé que celui des globulines , la relation est inverse chez les animaux de la ferme . Chez les mammifères adultes , le taux global des protéines plasmatiques varie entre 6 et 8 %. La teneur du sérum en protéines totales diminue en cas d’alimentation carencée en protides . La fraction albumine diminue surtout dans les affections hépatiques car ces protides sont en grande partie synthétisés dans cet organe .Les globulines sont élaborées dans le foie et dans les cellules du système réticulo-endothélial ; il y’ a augmentation dans beaucoup de maladies infectieuses chroniques . Le plasma sanguin est par définition un liquide intra-vasculaire et la pression osmotique ( ou oncotique ) intra-vasculaire générée par les protéines plasmatiques s’oppose à la pression hydrostatique dans les espaces tissulaires .Cette action empêche la sortie de trop grandes quantités de liquides plasmatiques dans la région des capillaires artériels ; dans les capillaires veineux d’autre part , la pression oncotique des colloïdes du plasma permet la réabsorption d’une partie de l’eau tissulaire. Lorsque le taux des protéines du plasma diminue (hypoprotéinémie), cette réabsorption est défectueuse et l’eau s’accumule en excès dans les tissus. L’hypoprotéinémie est rencontrée dans diverses infections chroniques accompagnées de troubles nutritionnels (comme dans la tuberculose, la cachexie , l’insuffisance hépatique , le syndrome néphrotique , les endoparasites ,les syndromes de malabsorption et de mal-digestion et dans le cas de brûlures sévères ). L’albumine est synthétisée dans le foie, elle est formée d’une seule chaîne comportant 610 acides aminés. Elle sert comme molécule de transport pour la bilirubine, les acides gras, les éléments à l’état de trace et plusieurs drogues. Certains de ses sites de liaison sont hautement spécifiques et saturables alors que d’autres le sont beaucoup moins (Kolb, 1975).Ce sont les sérums-albumines qui jouent le rôle principal dans le maintien de la pression oncotique du plasma car elles ont un poids moléculaire plus faible que celui des globulines. La teneur globale en protéines du sérum est en relation avec celle du secteur hydrique , aussi le taux de protéines sériques semble augmenter ou à l’inverse diminuer en cas de déshydratation ou d’hyperhydratation . La teneur en albumine du sérum diminue chez les vaches laitières à la période du vêlage et ne retrouve sa valeur du départ que progressivement au cours des trois mois suivants (corrélation directe avec la lactation ).En revanche , le bilan protidique n’est que très peu influencé par la méthode d’élevage et d’alimentation (Rosenberger, 1979 ). Tableau n° 16 : Normes physiologiques des protéines totales et de l’albumine sériques chez le mouton. 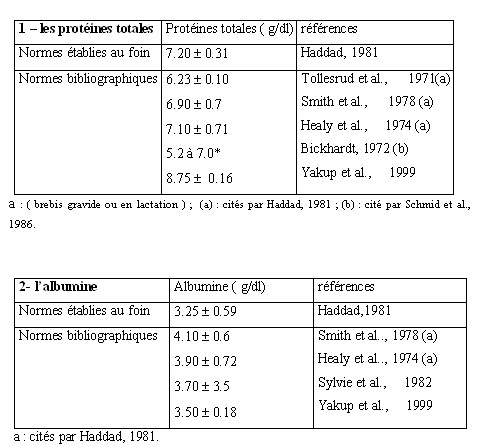 Il est cependant reconnu qu’une ration pauvre en protéines engendre une diminution de la production laitière et du contenu protéique du lait .Les résultats de Paquay , Godeau , De Baere et Lousse (cités par Payne 1983 ) suggèrent que les rations contenant moins de 15 % de protéines brutes dans la matière sèche réduisent la production de lait chez les vaches hautes productrices . Chez les autres catégories d’animaux , une balance protéique négative entraîne sans aucun doute une baisse des protéines sanguines et une perte de protéines musculaires. Vraisemblablement , la baisse de la synthèse des protéines peut agir sur la production des hormones protéiques comme la FSH et la LH dans l’hypophyse et affecter la fertilité. Rowlands , Little et Kitchenham cités par Payne (1983 ) ont trouvé une corrélation entre le taux d’albumine sérique et la fertilité chez les vaches laitières . Le mini-profil biochimique de Blowey cité par Haddad 1981, portant sur 03 paramètres, glucose , urée et albumine fait ressortir que : - le taux de glucose indique le niveau énergétique de la ration - le taux d’urée reflète la quantité d’ammoniac dans le rumen - le taux d’albumine témoigne du stockage des protéines Dans le cas où l’on trouve , un taux de glucose faible associé à un taux d’urée élevé et un taux d’albumine presque normal cela indique que le troupeau reçoit un foin de mauvaise qualité. Beaucoup de chercheurs , ont noté un faible taux d’albumine et des œdèmes chez les veaux et les vaches qui ingèrent une ration protéique très pauvre .Dans ce cas ,le changement dans la concentration en albumine est dû à l’insuffisance des entrées. Une autre cause d’hypo-albuminémie est l’augmentation non compensée des sorties. - Facteurs de variations Les augmentations et les diminutions de la teneur globale en protéines dans le sérum sanguin , d’origine pathologique ,s’accompagnent également de modifications dans la répartition des diverses fractions protéiques du sérum. Ainsi chez les veaux et les jeunes bovins, on observe une hypoprotéinémie traduisant une hypo-albuminémie ,à la suite d’une diarrhée importante ou d’helminthoses gastro-intestinales par élimination de grandes quantités de protéines au niveau de la caillette ou de l’intestin .Bien que l’on ne sache pas avec certitude où se trouvent les sites de dégradation de l’albumine , il existe des maladies du foie et du tractus digestif comme la fasciolose et la maladie de Johne (ou la paratuberculose) qui entraînent une fuite intestinale excessive des proteines sériques dont le taux dépasse la capacité de synthèse hépatique . *Dans la fasciolose ovine la demi-vie de l’albumine passe de 400- 470 heures (valeurs normales) à 110- 280 heures. Cela est dû à la fuite de l’albumine, par l’intermédiaire de la bile, dans le tractus gastro-intestinal .( Payne 1983) *Dans la maladie de Johne , il semble que la synthèse protéique hépatique soit au contraire augmentée et il paraît évident que l’hypo-albuminémie a pour origine à la fois une augmentation des pertes et une diminution de l’absorption à travers la muqueuse endommagée de l’intestin grêle. Les moutons touchés par cette maladie perdent en moyenne 4 g de protéines par jour . *Chez les vaches laitières, les hypo-protéinémies sont la plupart du temps en relation avec une anémie marquée .Elles sont le symptôme corrélatif d’une affection hépatique (troubles de la synthèse hépatique des protéines ) ou d’une affection rénale (protéinurie ). Chez les malades atteints de néphrose amyloïde, l’hypoprotéinémie se développe rapidement et résulte de la perte simultanée de protéines au niveau des reins (urines )et de l’intestin (excréments). L’augmentation de la teneur globale en protéines dans le sérum (hyperprotéinémie ) est liée chez les bovins ,à une hyperglobulinémie gamma ,reflet en général d’un processus pathologique grave ,aigu ou chronique ,inflammatoire, purulent ou pyohémique, métastatique dont le siège devra être déterminé à l’aide d’autres méthodes d’examen. VI.2.2.2- Urée plasmatique L’urée est le résidu principal de la dégradation des protéines par l’intermédiaire de l’acide glutamique et de la glutamine , du carbamyl-phosphate et du cycle catalytique des acides aminés basiques (arginine .ornithine citruline). L’urée est synthétisée essentiellement dans le foie. Chez les ruminants, l’urée sanguine a une signification un peu différente des monogastriques . Chez les non-ruminants, l'urée produite dans le corps est toujours entièrement perdue dans les urines. Elle dépend non seulement de la composition du mélange des acides aminés qui passe du tube digestif dans le sang et du taux azoté mais aussi des facteurs qui permettent une plus ou moins bonne utilisation de l’ammoniac du rumen pour la synthèse des protéines bactériennes (Haddad 1981). A cet effet beaucoup d’études ont prouvé le passage de l’urée du sang vers le milieu ruminal , ce processus est très bénéfique pour le ruminant car les bactéries présentes dans le rumen-réseau sont capables d’utiliser l’azote apporté par l’urée pour la synthèse protéique (Rémond et al., 1996 ). les ruminants possèdent un mécanisme pour conserver l'azote lorsque leur ration est déficiente en azote. L'urée est le produit final du métabolisme des protéines dans le corps et elle est normalement sécrétée dans les urines. Chez les non-ruminants, l'urée produite dans le corps est toujours entièrement perdue dans les urines. Plusieurs auteurs estiment que l’urée sanguine est l’indicateur essentiel du taux azoté de la ration . Elle augmente : • avec l’importance des apports azotés • avec un catabolisme accru par le jeûne (Ndibualonji et al., 1997 ) • suite à une intoxication par l’urée lors d’adjonction dans la ration , • suite à une sous nutrition énergétique Par contre un taux faible de l’urée sanguine peut signifier que la ration est riche en amidon ou faible en apport azoté . (Haddad 1981, Rémond et al.,1996). Cependant Wolter (1992) estime que c’est l’urée du lait qui constitue un bon indicateur du rationnement azoté . - Facteurs de variations L’urémie n’est pas influencée par la génétique mais peut avoir des variations suivant : - la race , l’âge aussi bien chez les ovins que chez les bovins (Haddad 1981). Tableau n°17 : Normes physiologiques de l’urée sanguine chez le mouton 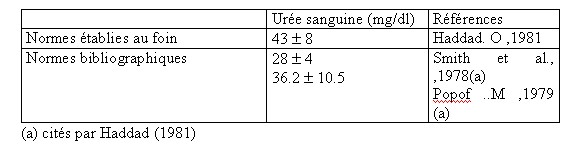 - la saison où le taux d’urée sanguine en été est plus élevé. Cette augmentation pourrait être due à la fertilisation des sols et l’addition de très grande quantités d’azote non protéique dans la ration (Savaria 1975 cité par Haddad, 1981 ). - la restriction de distribution d’eau s’accompagne d’une augmentation de l’urémie . - la gestation n’a pas d’effet sur l’urémie , mais elle augmente au cours du premier mois de lactation. - En pathologie , l’urémie s’observe le plus souvent dans les affections rénales . VI.2.2.3– CréatinineLa créatine en se déshydratant spontanément donne naissance à la créatinine. La créatinine est une petite molécule cyclique dont le taux plasmatique , pratiquement indépendant de l’apport protéique alimentaire reflète la masse musculaire du sujet concerné. Pour un sujet donné , le taux plasmatique , la quantité de créatinine éliminée quotidiennement dans les urines , constituent des paramètres biologiques remarquablement fixes. Le taux de la créatinine est indépendant de l’alimentation, il augmente pendant la deuxième moitié de la gestation et se trouve en corrélation négative avec la production laitière après le vêlage. On constate des augmentations pathologiques de ce paramètre en particulier lors d’affections rénales graves ou d’entérite. | |
| | | | merouane
Messages : 2
Points : 2
| | Sujet: Re: COURS DE NUTRITION ANIMALE DEUXIEME ANNEE (Par DR MEZIANE TOUFIK) Jeu 12 Nov 2009 - 12:45 | |
| coucou amira.tu m'as rapellé l'enseignant d'alimentation et les belles années à l'université. j'éspère lire de toi encore et encore.merci | |
| | | | mazagan
Messages : 30
Points : 37
| | Sujet: Re: COURS DE NUTRITION ANIMALE DEUXIEME ANNEE (Par DR MEZIANE TOUFIK) Mer 17 Mar 2010 - 1:07 | |
| | |
| | | | laadjal

Messages : 54
Points : 34
| | Sujet: Re: COURS DE NUTRITION ANIMALE DEUXIEME ANNEE (Par DR MEZIANE TOUFIK) Mar 3 Aoû 2010 - 11:08 | |
| | |
| | | | Dakhi_55
Messages : 8
Points : 8
| | Sujet: Re: COURS DE NUTRITION ANIMALE DEUXIEME ANNEE (Par DR MEZIANE TOUFIK) Ven 6 Aoû 2010 - 14:19 | |
|  merci | |
| | | | 3malik1

Sexe : 
Messages : 2
Points : 2
 Date de naissance : 13/11/1972 Date de naissance : 13/11/1972
 Age : 51 Age : 51
 Emploi : agronome Emploi : agronome
| | Sujet: Re: COURS DE NUTRITION ANIMALE DEUXIEME ANNEE (Par DR MEZIANE TOUFIK) Sam 15 Oct 2011 - 20:46 | |
| je suis revenu 20 ans en arriére. Avec vous je possede encore ma carte d'étudiant. Merci beaucoup et bon courage pour le reste. Merci! | |
| | | | sonia fre

Sexe : 
Messages : 1
Points : 1
Date de naissance : 14/09/1994
Age : 30
Emploi : veterinaire
| | Sujet: Re: COURS DE NUTRITION ANIMALE DEUXIEME ANNEE (Par DR MEZIANE TOUFIK) Sam 1 Mar 2014 - 21:31 | |
| merci bcp
chui en periode des exams et vous m'avez bcp aidé
merci encore une fois | |
| | | | taibivet
Sexe :
Messages : 1
Points : 1
Date de naissance : 25/10/1988
Age : 35
Emploi : Docteur vétérinaire
| | Sujet: Re: COURS DE NUTRITION ANIMALE DEUXIEME ANNEE (Par DR MEZIANE TOUFIK) Dim 15 Oct 2017 - 19:35 | |
| cours d'alimentation animale 2eme année vét svp .merci | |
| | | | nesrine hadj
Sexe :
Messages : 1
Points : 1
Date de naissance : 01/03/1989
Age : 35
Emploi : ETUDIANTE MASTER 1 PRODUCTION ANIMALE
| | Sujet: Re: COURS DE NUTRITION ANIMALE DEUXIEME ANNEE (Par DR MEZIANE TOUFIK) Sam 29 Déc 2018 - 19:53 | |
| Slm merci beaucoup pour votre effort j'ai besoin des examens de cette spécialité et ce module svp | |
| | | | Contenu sponsorisé
| | Sujet: Re: COURS DE NUTRITION ANIMALE DEUXIEME ANNEE (Par DR MEZIANE TOUFIK) | |
| |
| | | | | | COURS DE NUTRITION ANIMALE DEUXIEME ANNEE (Par DR MEZIANE TOUFIK) | |
|
| | Permission de ce forum: | Vous ne pouvez pas répondre aux sujets dans ce forum
| |
| |
| | Site partenaire |  |
|
