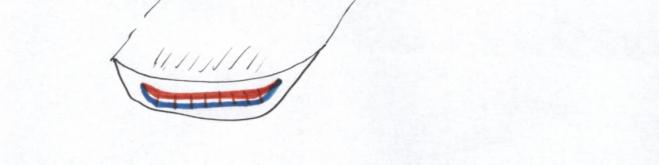
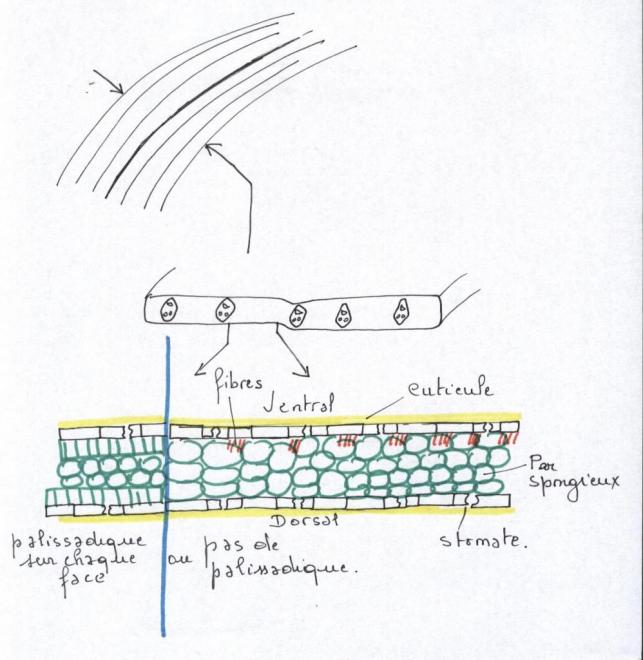
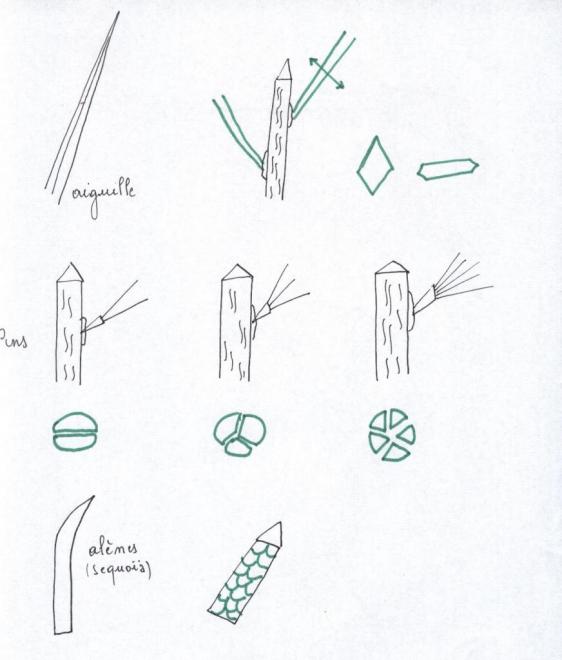
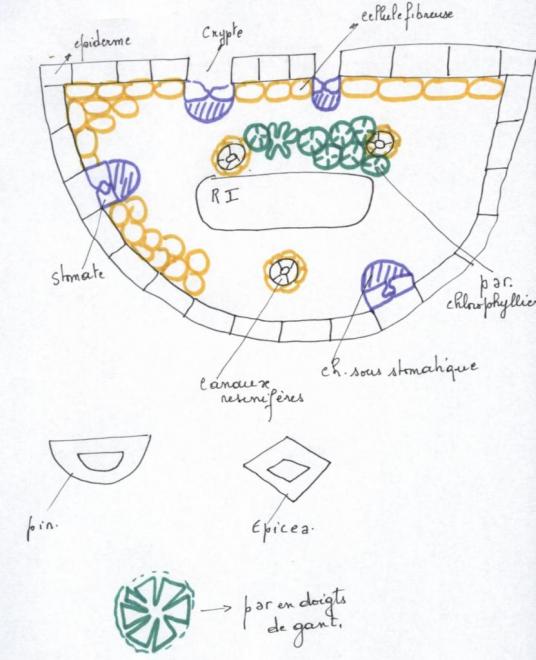
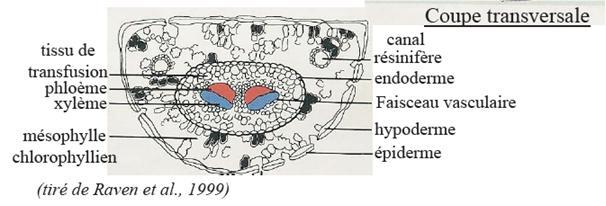
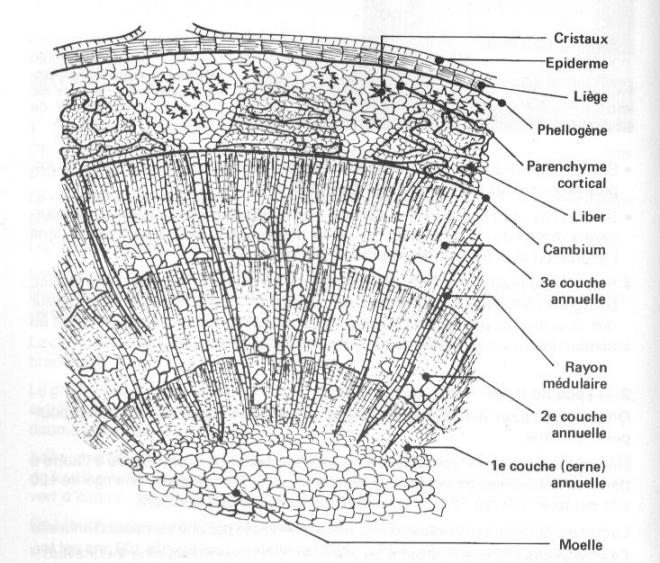
VI.5 LA STRUCTURE ANATOMIQUE SECONDAIREC’est l’ensemble des tissus qui contribuent à l’épaississement de la plante
JUSTIFICATIONLes méristèmes situés à l’extrémité des
rameaux s’accroissent – du moins théoriquement – d’une manière indéfinie
(plantes pérennes).
La plante pérenne doit grandir pour
vivre : elle produit chaque année de nouvelles feuilles, c'est-à-dire
des nœuds ; les feuilles sont séparées par des intervalles ou
entre-nœuds, d’où élongation.
Nous verrons plus tard que la plante -et
plus particulièrement le type arbre- grandit en hauteur : tiges à cime
pointue ; ultérieurement, la cime s’arrondit parce que l’élongation
porte surtout sur les rameaux latéraux.
De l’apparition de nouvelles feuilles
résulte une augmentation constante de conducteur ; la plante à besoin
d’un soutien au fur et à mesure qu’elle se développe : la tige
s’épaissit.
Ce soutien est réalisé par la structure
secondaire grâce à l’activité cambiale. Le cambium est le premier
méristème secondaire de la tige :
il donne à
la fois du tissus conducteur et du soutien.REMARQUE : un palmier, plante monocotylée, garde un nombre de feuilles
constant : les feuilles les plus vieilles tombent lorsque les nouvelles
feuilles apparaissent. C’est pourquoi, il n’est pas astreint à
s’épaissir, et sa tige (stipe) conserve une épaisseur constante.
L’épaississement de la tige provoque
l’éclatement de l’épiderme. Afin de suppléer à la protection déficiente
de l’épiderme , un protecteur secondaire apparaît sous la forme d’un
tissus subérisé appelé
liègeLe liège provient d’un méristème particulier , sous-jacent à l’épiderme : c’est le
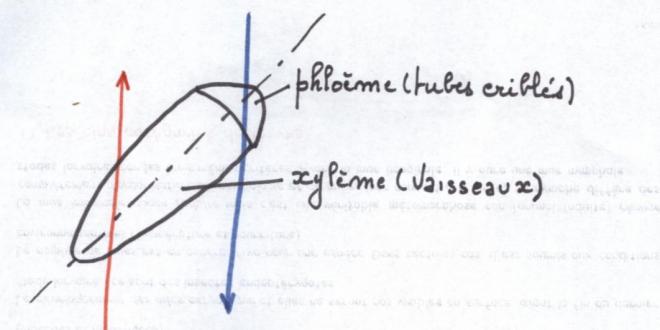
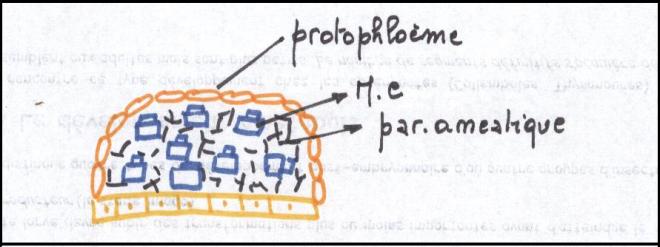
phellogèneLA STRUCTURE SECONDAIRE CONDUCTRICE DE LA TIGECette structure s’adresse à toute les plantes possédant du cambium, c'est-à-dire les Dicotylées et les Gymnospermes ;
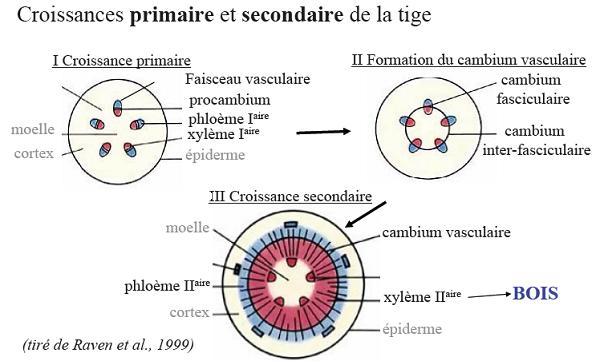
Les files radiales de cellules
méristématiques subissent progressivement une différenciation qui
commence avec les cellules les plus agées. Les cellules internes
deviennent des vaisseaux ponctués ; les cellules externes évolueront
progressivement en tubes criblés et cellules compagnes.
Pendant le même temps, le cambium fait
apparaître de nouvelles cellules qui refoulent les précédentes et
« ouvrent » le faisceau progressivement puisque le
xylème et le phloème primaires s’écartent de plus en, plus.
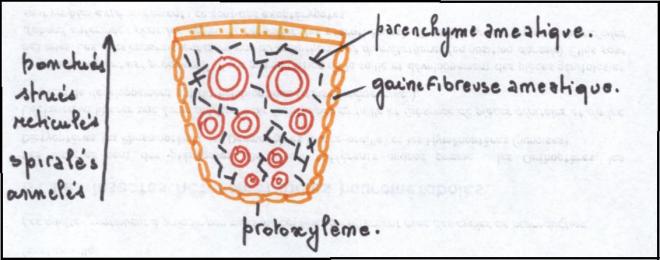
Il faut noter que les cellules issues du
cambium comprennent non seulement du conducteur , mais aussi du
parenchyme et du soutien. Le parenchyme produit est à l’origine des
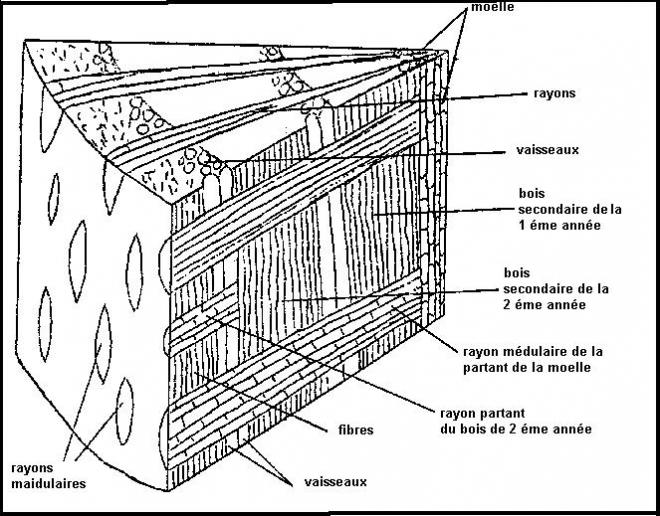
rayons médullaires ou
rayons ligneux orientés radialement et composé de cellules parallélépipédiques, Ces
rayons constituent le système radial par opposition au système axial
constitué des tissus conducteurs secondaires
REMARQUESLes vaisseaux produits par le cambium sont du type ponctués ( résistance à l’écrasement).
Chez les Angiospermes, les tubes criblés sont flanqués d’une cellule compagne, par division latérale de la cellule originelle.
L’ensemble des vaisseaux ponctués, du
parenchyme et des fibres produits par le cambium du côté xylème
constitue un nouveau tissus
se développant de manière centrifuge et est appelé
bois ou xylème secondaireL’ensemble des tubes criblés issus du
cambium , avec les cellules compagnes, les fibres et le
parenchyme appartient à un nouveau tissus conducteur , progressant de
manière centripète et est appelé
liber ou phloème secondaire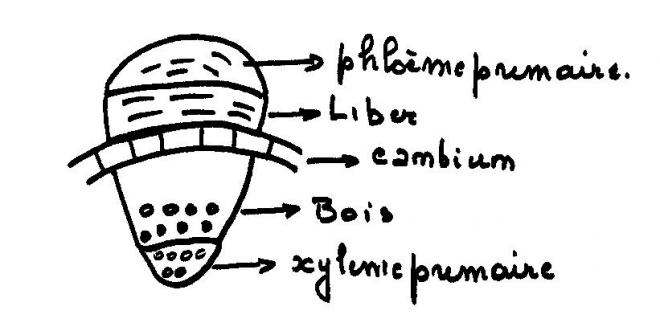 1.Le cylindre cambial
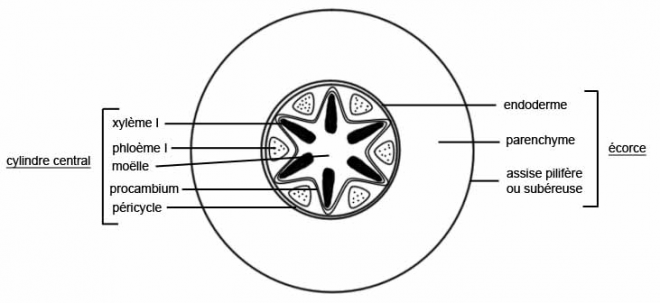
1.Le cylindre cambialReprenons les trois types de tiges étudiées en structure primaire
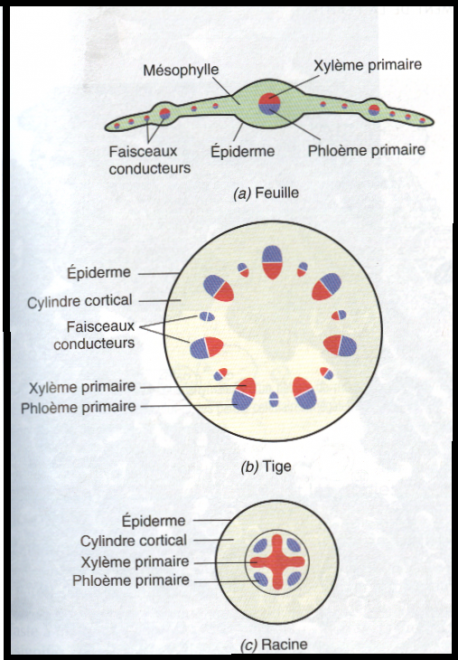
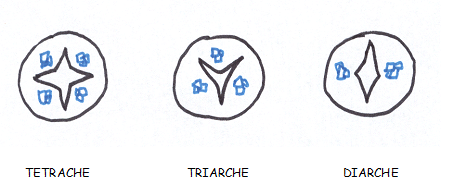
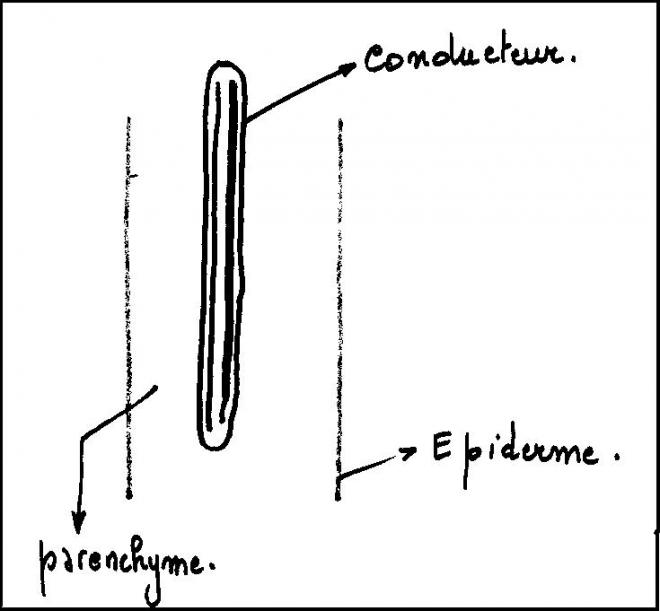
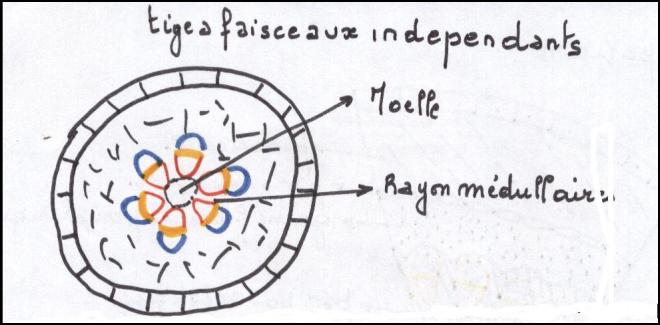
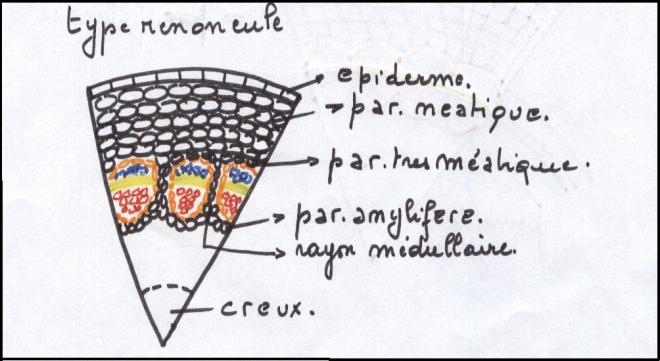
Le type herbacé ou renonculeLes faisceaux sont peu nombreux et espacés les uns des autres dans un cercle inscrit à l’épiderme
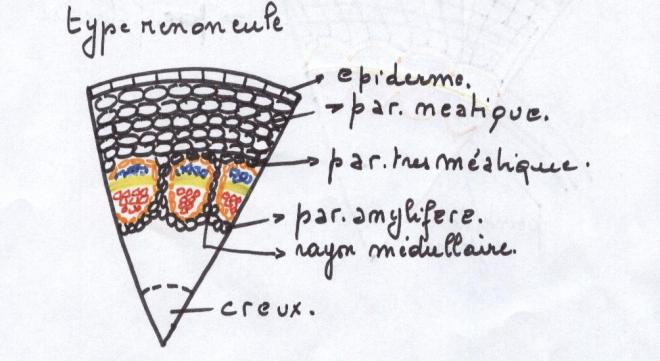
Chaque faisceau connaît des divisions
cambiales, mais celles-ci sont rapidement limitées dans le temps , car
les cellules parenchymateuses des rayons
médullaires sont très évoluées, méatiques, et d’autre part les rayons médullaires sont larges.
C’est pourquoi le cambium fonctionnera aussi longtemps que le permettra l’élasticité des rayons médullaires
En raison de la largeur des rayons
médullaires et de la profonde différenciation de leurscellules , de
telles plantes sont incapables d’épaissir leur tige , sinon d’une
manière très limitée.
Elles sont donc vouées à rester de petite taille parce que
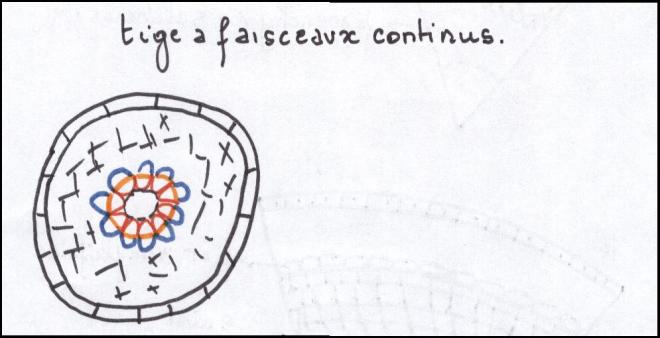
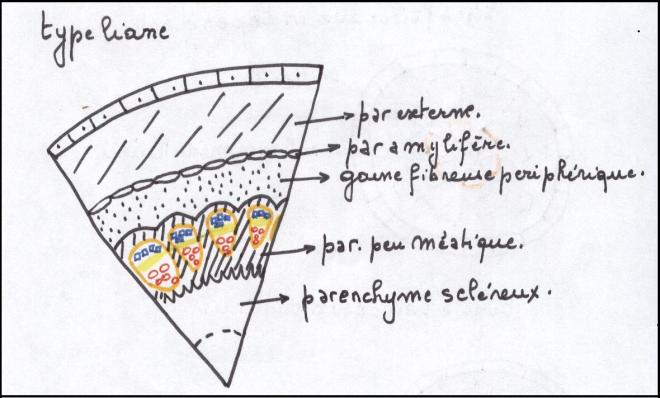
les plantes herbacées n’ont pas de cylindre cambialLe type liane- Les faisceaux sont beaucoup plus
nombreux et par conséquent, les rayons médullaires nettement plus
étroits que dans le cas précédent.
- Les cellules parenchymateuses des
rayons médullaires situées au niveau du cambium sont petites, peu ou pas
vacuolisées. Elles sont capables de subir des divisions cellulaires
- Dans ce type de tiges, le
fonctionnement du cambium entraîne dans ses divisions celles du
parenchyme qui lui est contigu dans le rayon médullaire et l’ensemble
fonctionne comme un
cylindre cambial continu mais hétérogène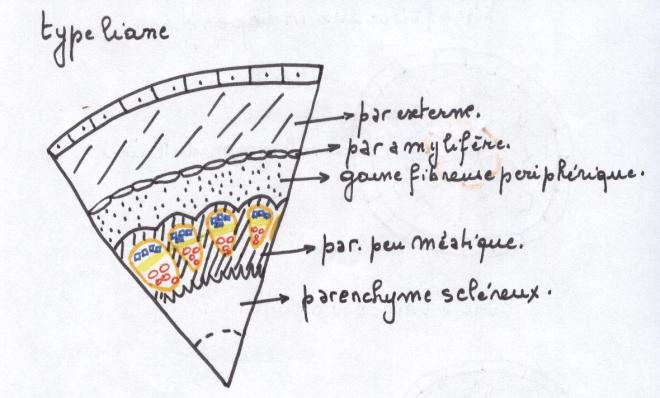
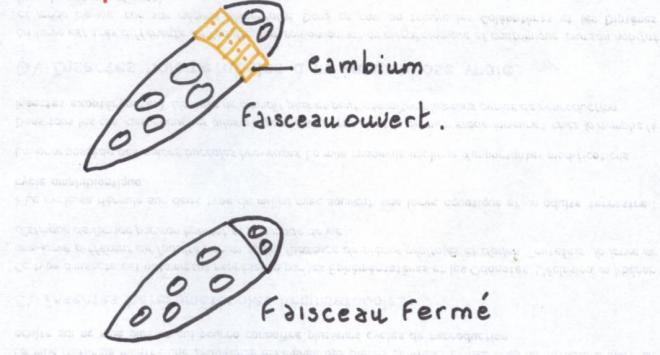
Hétérogène car constitué par du
cambium vrai ou intrafasciculaire ( = à l’intérieur des faisceaux ) , producteur de bois et de liber et par du
cambium
faux ou extrafasciculaire qui est du parenchyme et ne produira que du parenchyme.
Une telle tige sera capable d’acquérir un certain développement ;
selon la proportion de cambium vrai qu’elle contient par rapport au
cambium faux , producteur de parenchyme.
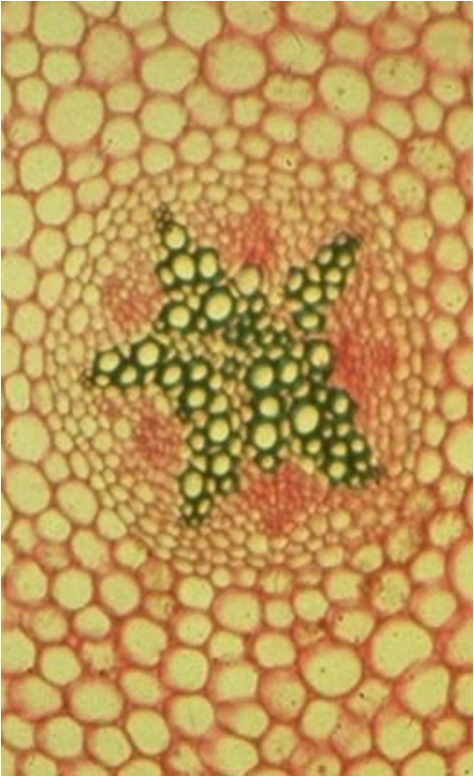
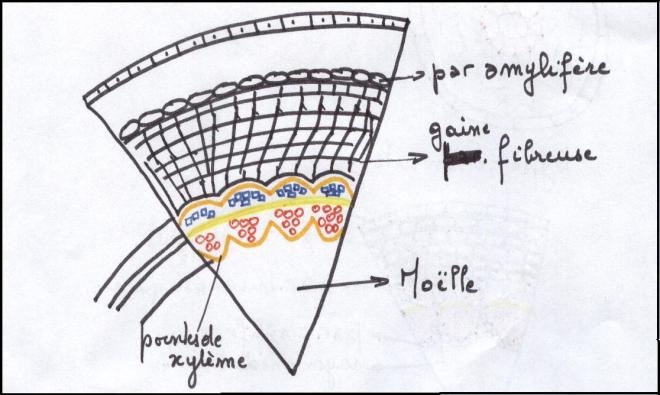
Le type arbreLes faisceaux sont séparés par des rayons médullaires très minces ,
au point que les faisceaux apparaissent comme contigus et ne se
distinguent que par
les pointes de protoxylème faisant saillie dans la moelle.
Le cambium vrai prédomine très largement sur le cambium faux.
On admet que le
cylindre cambial est homogèneCe troisième type de tiges est capable de produire des tiges importantes à base de bois. 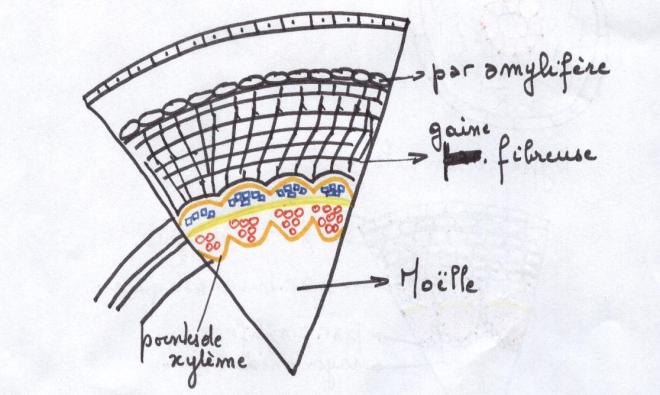 CONCLUSIONS
CONCLUSIONS1. Une plante de type herbacée est dépourvue d’anneau cambial , elle est condamnée à garder une petite taille.
2. Une plante de type liane est dotée d’un
cylindre cambial hétérogène . Ce cylindre hétérogène contient une trop grande proportion de parenchyme par
rapport
à son cambium vrai et ultérieurement à son bois : elle est incapable de
former un tronc assez rigide pour supporter le poids d’une cime. Elle
connaîtra néanmoins un certain
développement.
3. Une plante du type arbre , au cylindre cambial dit homogène , possède assez de bois pour supporter une cime importante.
La production du cylindre cambial homogèneOn peut résumer comme suit les divers tissus produits par le cambium
1. Production du cambium vrai ou intrafasciculaire1 .1 Intérieur de la tige = bois1.1.1
Dicotylées ligneuses ou feuillus- Trachées ( vaisseaux ponctués)
- Parenchyme ligneux
- Fibres
- Eventuellement : sécrétion (gommes)
1.1.2
Gymnospermes ou résineux- Trachéides ( vaisseaux aréolés)
- Parenchyme ligneux
- pas de fibres
- Canaux résinifères
1.2 Extérieur de la tige = liber- Tubes criblés plus cellules compagnes
- parenchyme libérien
- fibres
2. Production du cambium faux ou extra fasciculaire 2.1 Rayons médullaires- Parenchyme
A partir d’une certaine épaisseur, les rayons médullaires donneront
la maille du bois.
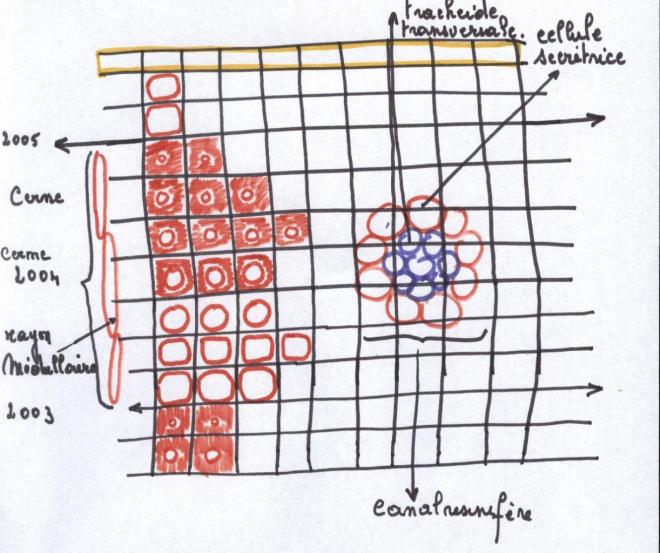
Périodicité de fonctionnement du cylindre centralElle est corrélative au climat général et à la période de végétation .
En régions tempérées : l’activité cambiale démarre au printemps et demeure active durant
l’été ; elle ralentit à l’automne et s’interrompt durant la période
hivernale , elle est donc saisonnière.
En régions tropicales :le fonctionnement cambial s’arrête en saison sèche et reprend en saison des pluies.
En régions équatoriales : Beaucoup d’essences ont un fonctionnement cambial permanent
On peut dire, en conclusion, que sauf en régions équatoriales,
l’activité cambiale est marquée par des temps d’arrêt. Ces temps
d’arrêt délimitent des cernes ou productions de bois périodiques
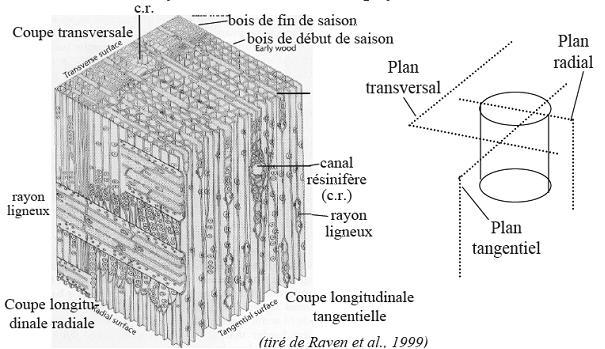
 3. Le bois des Gymnospermes
3. Le bois des GymnospermesIl est à base de trachéides qui apparaissent en séries radiales
telles qu’elles sont issues du cambium. Elles ont pratiquement toutes le
même diamètre
extérieurLes trachéides issues du cambium juste avant le repos de végétation
sont à lumen étroit parce que la membrane secondaire est importante :
elles jouent
surtout un rôle de soutien - donc de fibres – puisque les trachéides sont entrecroisées à la manière de fibres.
Les trachéides formées en début de période de végétation offrent au
contraire un lumen large , des parois secondaires indiquant une faible
lignification :
elles ont surtout une fonction vasculaire donc conductrice car à ce moment il faut à la plante beaucoup de sève minérale.
Les rayons médullaires sont étroits, ayant l’épaisseur d’une cellule.
 Conclusion
Conclusion : un cerne ou couronnecirculaire de bois est constitué par du
bois initial, poreux et clair à fonctions surtout conductrices, ensuite à du
bois final peu
poreux et foncé , à fonction surtout de soutien et l’ensemble
constitue un cerne qui est surtout matérialisé par sa transition avec le
bois des années contigues
En résumé : Ce bois
est constitué - outre les canaux résinifères et le parenchyme- par des
cellules du type vaisseaux fermés ou trachéides, formant des séries
radiales et de
largeur hors tout constante.
Seul varie la largeur du lumen qui fait de ces cellules en ordre
principal du conducteur ou du soutien selon l’époque de l’année.
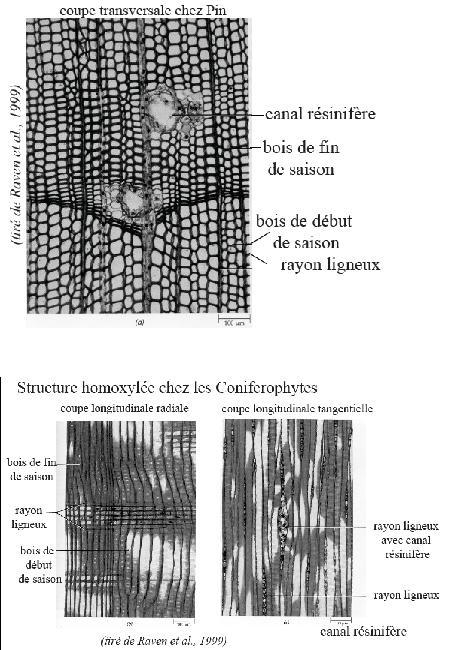 Remarquesc omplémentaires
Remarquesc omplémentaires- La régularité de la disposition cellulaire donne au bois produit un
« fil » droit, notion importante en technologie où cette particularité
est appréciée.
- Les rayons médullaires limités à la largeur d’une cellule (
parenchyme) font que le bois des « résineux » est dépourvu de maille.
- Les résineux ont un bois dit hétérogène ( voir ci après), l’élément
constant dans une cerne est le
bois final.
- Le bois initial est composé de cellules à lumen très large, mais sans que soit modifié le périmètre extérieur de ces cellules
Ce sont des plantes tropophylles, c'est-à-dire à feuilles caduques.
Au départ, le cambium engendre des cellules en files radiales
Mais au moment de la reprise de la végétation, donc au printemps en
régions tempérées, l’arbre doit refaire son appareil foliaire dans son
entièreté. Aussi la plante forme-t-elle des trachées (vaisseaux ouverts
de type ponctué) à lumen larges , cloisons peu épaisses : c’est le bois
initial à fonctions conductrices
dominantes, à tel point que la disposition radiale au départ du cambium est bousculée.
VI.Le bois des Dicotylées ligneuses ou feuillusCe sont des plantes tropophylles, c'est-à-dire à feuilles caduques.
Au départ, le cambium engendre des cellules en files radiales
Mais au moment de la reprise de la végétation, donc au printemps en
régions tempérées,l’arbre doit refaire son appareil foliaire dans son
entièreté. Aussi la plante forme-t-elle des trachées (vaisseaux ouverts
de type ponctué) à lumen larges , cloisons peu épaisses : c’est le bois
initial à fonctions conductrices
dominantes, à tel point que la disposition radiale au départ du cambium est bousculée.
Par la suite , ce sont les fonctions de soutien qui l’emportent : les
trachées sont moins larges , moins nombreuses et les fibres acquièrent
une proportion de
plus en plus importante alors qu’elles existaient en proportions réduite dans le bois initial.
En fin de végétation, il n’existe quasiment plus que des fibres dans
le cerne, toutefois certaines espèces produisent un anneau de parenchyme
en fin d’accroissement annuel.
Au bois initial clair et poreux, succède le bois final, foncé,peu poreux et résistant.
L’année suivante, le même cycle se répète et le bois initial succède
sans transition au bois final de l’année précédente. Au contraire ,
le
bois initial et le bois final d’une même année de croissance présente
un dégradé progressif de porosité, de teinte , de composition et
d’aspect.
VII Notions de bois hétérogènes et de bois homogènesUn bois est dit hétérogène
lorsque son bois initial est très poreux par rapport au bois final ;
les gros vaisseaux du bois initial sont visibles à l’œil nu en section transversale ( cas des résineux déjà cités, du chêne du frêne, de l’orme …)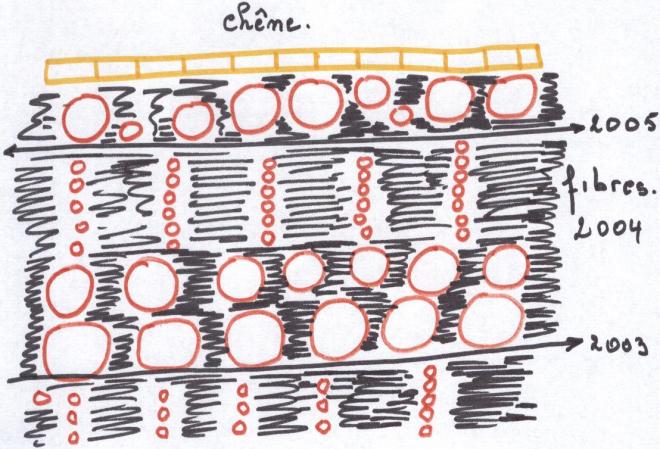
Un bois est dit
homogène lorsque la distinction précédente n’est pas possible à l’oeuil nu ( cas du hêtre, du peuplier, du bouleau etc…)
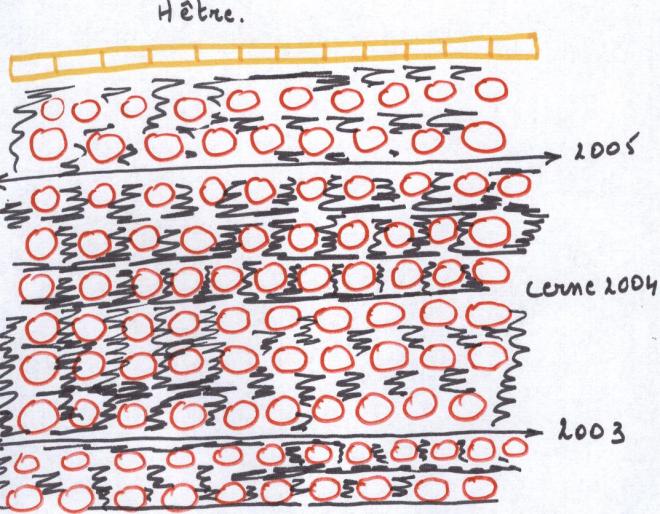
Le bois de tous les résineux est hétérogène ; les feuillus ont des bois soit
homogènes, soit hétérogènes.
La distinction entre le bois d’un feuillu et celui d’un résineux se
ramène à la reconnaissance de deux bois hétérogènes dont il faut
rechercher
l’élément constant au travers des cernes , L’élément constant chez les résineux est le bois final ; l’élément constant chez les feuillus est le bois initial,
V.I.I.I
L’ajustement du cylindre cambialEn l’an 2010 , le secteur de tige délimité par le schéma ci-dessous
sera élargi considérablement. Pour un même angle au centre, l’arc
cambial correspondant
sera plus développé
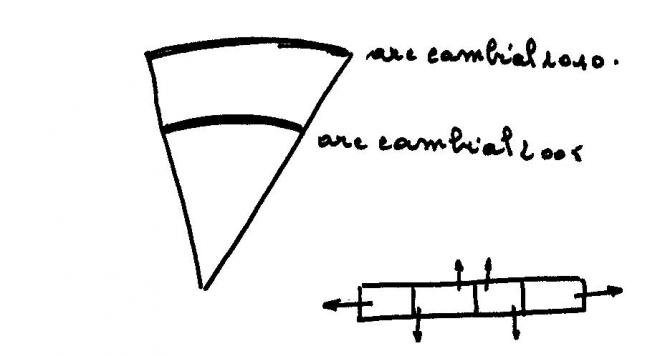
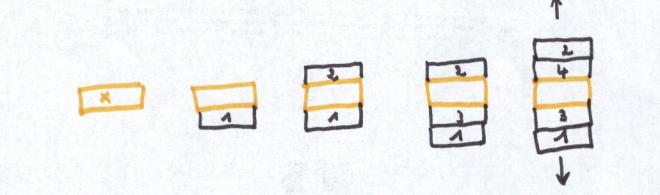
D’une année à l’autre, l’arc cambial recule vers l’extérieur de la
tige tout en s’appuyant sur le dernier cerne qui vient d’être formé.
Les divisions cambiales, surtout abondantes dans le sens radial(vers
l’intérieur puis vers l’extérieur ) réaliseront également des
cloisonnement latéraux ( = dans le sens tangentiel)en nombre limité et
dans les limites compatibles avec les nécessités : c’est l’ajustement du
cylindre cambial.
IX.
La production de liberElle est toujours moins importante que celle de bois , car il faut moins de sève élaborée que de sève brute.
Il n’existe pas de zonation dans le liber comme il en existe dans le bois

Comme on peut le voir sur le schéma précédent, les massifs libériens
se présentent en trapèzes ( section transversale) avec la petite base
vers l’extérieur de la tige tandis que la grande base s’appuie sur le
cambium
Le liber se développe de manière centripète
L’appellation de liber ou phloème secondaire vient de ce que les
tissus constituant le liber sont progressivement comprimés entre le bois
( à l’intérieur de la tige ) et le liège ( à l’extérieur de la tige) .
D’où l’aspect en livre vu par la tranche (latin : liber). Seules
fonctionnent les assises le plus internes du
liber ; elles sont aussi les plus récentes.
Pour rappel, il faut souligner que le liber est composé outres les
tubes criblés+ cellules compagnes, de parenchyme et surtout de fibres.
Celles-ci sont plus ou
moins nombreuses suivant les espèces.
IX.
Notion d’aubier et de duramenLa sève alimentaire circule par le liber. Etant donné le mode de
formation centrifuge du bois, le liber devient de plus en plus éloigné
des parties les plus internes du bois Certes, les vaisseaux ( trachées
ou trachéides) ainsi que les fibres sont des cellules mortes, mais le
bois contient également des cellules vivantes : le parenchyme ligneux.
Le parenchyme ligneux de moins en moins alimenté en profondeur
meurt.D’autre part, les vaisseaux des cernes les plus anciens perdent
leurs fonctions conductrices.
En général, les 12-15 cernes annuels les plus externes d’une tige
gardent leur parenchyme vivant et leurs vaisseaux fonctionnels.
Dans une tige ligneuse âgée, on distingue :
- Vers l’intérieur : le
bois de cœur ou duramen = bois non fonctionnel- Vers l’extérieur : en moyenne les 12-15 cernes les plus externes =
bois fonctionnel ou aubierL’aubier, seul
assure la conduction de la sève montante : ce bois est humide, bourré de
réserves ( amidon) ; il se prête idéalement aux attaques fongiques ou
entomologiques.
Si l’aubier est indispensable aux fonctions physiologiques de la tige
(conduction), il est à rejeter du point de vue technologique.
Le duramen est au contraire , un bois qui a cessé ses fonctions
conductrices , il est sec et duraminisé et, de ce fait, le seul apprécié
en technologie.
La duraminisation est une succession de stades physiologiques qui
transforment les cellules ligneuses du stade bois fonctionnel au stade
non fonctionnel ce qui revient à une mort physiologique du bois.
La duraminisation se manifeste en plusieurs stades.
Le parenchyme ligneux meurt après avoir produit des thyles, sortes de
hernies qui pénètrent dans les vaisseaux à la faveur des ponctuations
et bouchent les
lumens des cellules conductrices.
 Le bois ainsi obturé devient sec Pour
Le bois ainsi obturé devient sec Pour
beaucoup d’espèces, la duraminisation s’arrête à ce stade dénommé
thyllose. On dit de tels bois qu’ils ne présentent pas de distinction
entre l’aubier et le duramen : cas typique du hêtre. Ces bois ne
peuvent, en principe, être utilisés à l’extérieur, sauf imprégnation
artificielle ( créosotage). La
duraminisation se continue chez d’autres espèces parl’imprégnation, au
niveau de leurs parois , des cellules ligneuses à l’aide de substances
colorantes diverses qui sont la caractéristique de certains bois (
rougeâtre chez les acajous, grisâtre chez les noyers, rosé chez les
merisiers, etc …) ; parfois il y a imprégnation par des matières
antiseptiques ( tannins chez les chênes, résines chez les conifères, …). Parfois les deux substances sont présentes.Il arrive aussi que certaines matières minérales ( silice) s’incorporent au bois ( certains bois tropicaux). Remarques1)
Les essences présentant une distinction aubier-duramen sont évidemment
les plus intéressantes et les plus recherchées, car elles donnent
généralement à leur bois d’excellentes propriétés technologiques. Elles sont toutefois moins appréciées pour certains usages ( pâte à papier…).2)
Le faux cœur. A l’abbatage , certaines espèces présentent une
coloration qui s’atténue avec le temps et paraît liée au coloris du
substrat où elles se sont développées. C, est le cas des peupliers
hybrides euraméricains.3)
La maille d’un bois est une figure décorative que l’on obtient par
débit d’un arbre dans un plan longitudinal radial. Elle s’observe chez
les essences à rayons médullaires larges, plus larges que les traits de
scie, c’est pourquoi la maille est très apparente chez le chêne, l’orme,
le hêtre, etcPar contre elle est absente chez les résineux dont les rayons médullaires ont la largeur d’une seule cellule. 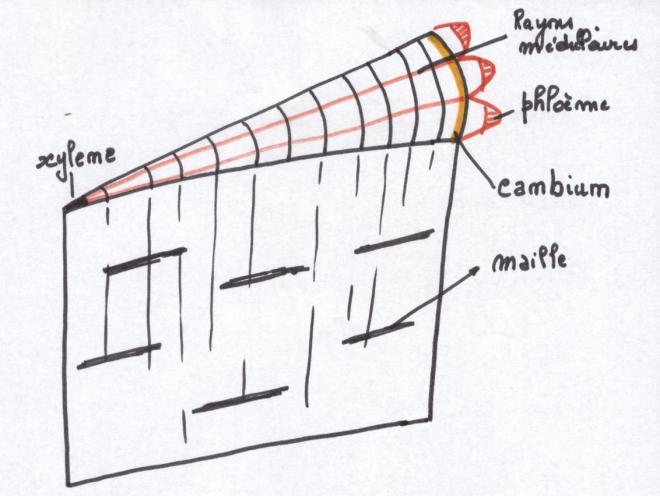
4)
Lesrayons parenchymateux internes se distinguent des rayons médullaires par leur origine.
Les rayons médullaires réunissent la moelle aux tissus extérieurs au cambium ; ils sont extra fasciculaires.
les rayons parenchymateux internes prennent naissance à partir de
cellules cambiales , à un endroit quelconque , ils sont
intrafasciculaires.
5) Le sciage sur boule, sur dosse, sur quartier :
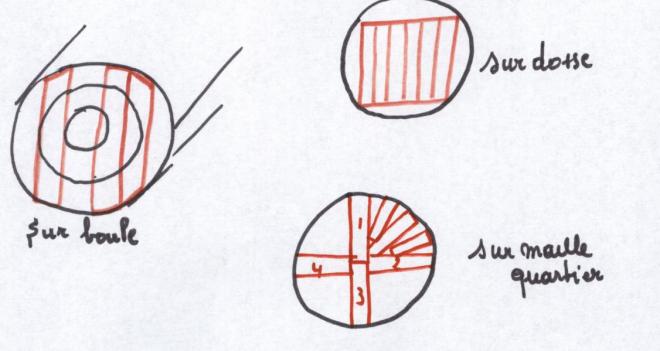
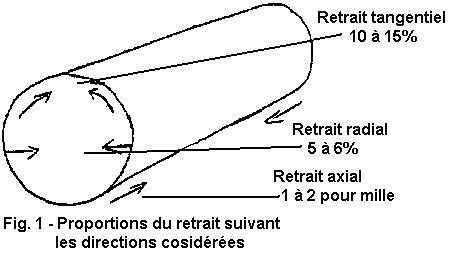
6) Le retrait du bois est la déformation que celui-ci subit au cours
de son séchage.L'importance des variations dimensionnelles n'est pas la
même suivant les espèces, mais est surtout variable suivant les
directions considérées :
1° - Dans le sens de l'axe de l'arbre le retrait est pratiquement négligeable.
2° - Dans le sens radial il est beaucoup plus important, atteignant jusqu'à 5 ou 6 % de la largeur des planches ou plateaux.
3° - Dans le sens tangentiel enfin le retrait peut atteindre
3 fois la valeur du retrait radial.
d)
Conséquences et palliatifs.
La rétractabilité du bois représente pour son utilisation rationnelle
un inconvénient grave. Pour le limiter il faut évidemment profiter au
maximum des directions dans les quelles le retrait est le moindre. Il
faut aussi tenir compte du fait que le retrait sur une face d'une pièce
de bois est d'autant plus important que cette face est plus éloignée du
cœur et que si les 2 faces
sont inégalement éloignées du cœur (ce qui
est le cas général), il en résultera une déformation, parfois même une
tendance à la fente).
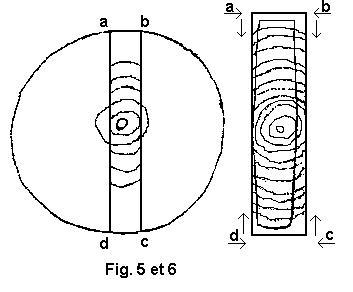
(voir figures 3 et 4 - 5 et 6 )
La pièce a b c d est dite débitée sur dosse.
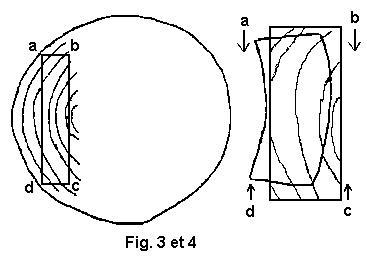
La
face a d, la plus éloignée du cœur subit un retrait plus important que
la face b c. Il en résulte une déformation qui détermine une convexité
de la face
la plus rapprochée du cœur.Les débiteurs disent que le bois "tire à cœur .
La pièce a b c d est dite débitée sur mailles ou sur quartier .La
face a d et la face b c sont également éloignées du cœur. Elles
subissent donc des retraits
tangentiels en a b et c d assez faibles
puisqu'ils se produisent sur l'épaisseur, sont égaux entre eux aussi. La
pièce subit une déformation beaucoup moins importante que la
précédente.
En conclusion :1° - Il faut essayer de mettre en oeuvre des bois qui auront été préalablement amenés à un
taux d'humidité en rapport avec les conditions d'utilisation.
2° - Chaque fois que possible on
préférera le débit sur quartier (ou un débit s'en approchant)
au débit sur dosse.
3° - Le séchage sera surveillé afin d'éviter les déformations brutales, génératrices des gerçures.
4° - L'emmagasinage des objets terminés se fera dans un local dont
l'atmosphère sera conditionnée en rapport avec l'utilisation (qui a été
prévue à la fabrication).
5° - Les enduits ( peintures, vernis, cires ),appliqués sur les
pièces s'opposent aux échanges d'humidité entre le bois et le milieu
ambiant. ( On peut de cette façon réaliser en bois des modèles de
fonderie absolument valable pour des pièces en métal. Le noyer est l'un
des bois les plus estimés pour la fabrication des modèles de mécanique.)
LA STRUCTURE SECONDAIRE DES PLANTES MONOCOTYLEESElle n’existe pas car ces plantes n’ont
pas de cambium et surtout
un anneau cambial n’est pas possible puisque les faisceaux sont dispersés à
toutes les profondeurs dans la tige.Ceci explique pourquoi les plantes n’épaississent pas leur tige.
Pourquoi les faisceaux sont-ils dispersés à l’intérieur de la tige ?
Les faisceaux foliaires sont nombreux car la nervation est
parallèle ; les faisceaux foliaires pénètrent dans la tige en réalisant
un parcours
oblique et se recourbent à des profondeurs et à des
distances diverses, puis se rattachent aux faisceaux préexistants après
un parcours qui les rapproche de la périphérie de la tige.C’est là
qu’ils sont généralement les plus densément représentés.
Le cas particulier des palmiers Ces plantes sont des monocotylées de grande taille mais ne sont pas des arbres.
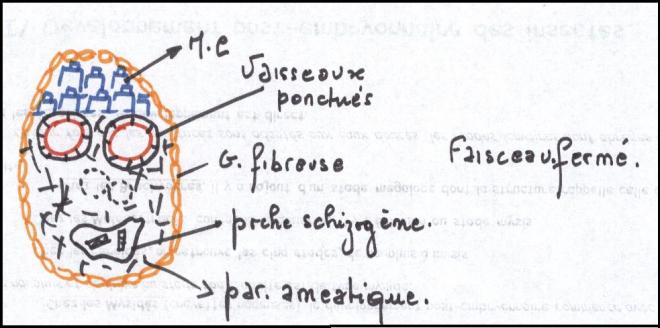
La tige du palmier porte le nom de stipe : elle est constituée de
faisceaux fermés , donc sans cambium ; ces faisceaux sont entourés de
fibres silicifiées et
les intervalles sont du parenchyme . Les palmiers ne sont pas des producteurs de bois
-
Le méristème des palmiers est particulier : Très développé, il peut avoir la largeur d’une assiette et présente la forme d’un dôme , les cellules sous
jacentes à ce méristème s’étirent latéralement ce qui est une exception
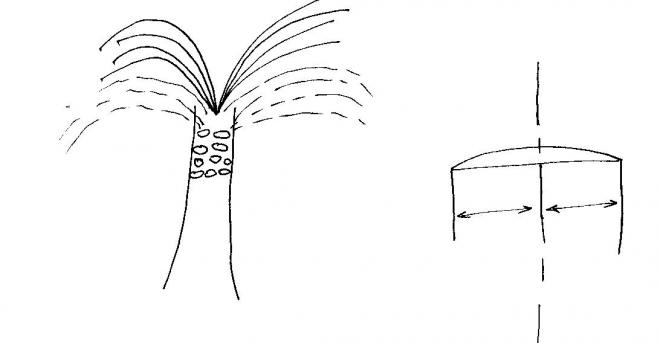
Ultérieurement, les cellules se lignifient et : elles deviennent
incapables de changer la forme et leur stipe devient cylindrique.
-
La disposition foliaire des palmiers. Pour rappel la phyllotaxie des palmiers est du type 8/21 : les feuilles sont ramassées en un vaste bouquet au
sommet de la tige (stipe).
Pourquoi ?
Parce que les entre nœuds sont très courts du fait de
l’étirement cellulaire déjà cité : les cellules sont allongées dans le
sens radial ; elles
sont étroites dans le sens longitudinal axial.
Le nombre de feuilles reste constant, les plus anciennes tombent lorsque les nouvelles se forment.
La base des feuilles demeure adhérente au stipe sous la forme d’un
chicot, ce qui donne un aspect particulier à la tige du palmier.
LA STRUCTURE SECONDAIRE PROTECTRICE DE LA TIGE
A l’intérieur de la tige, une pression s’exerce à la suite de l’action centrifuge du bois.
Il va de sois que l’épiderme unicellulaire ne suffit plus à assurer
la protection de la tige. La plante prévient ce risque et réalise un
méristème secondaire sous
épidermique de protection
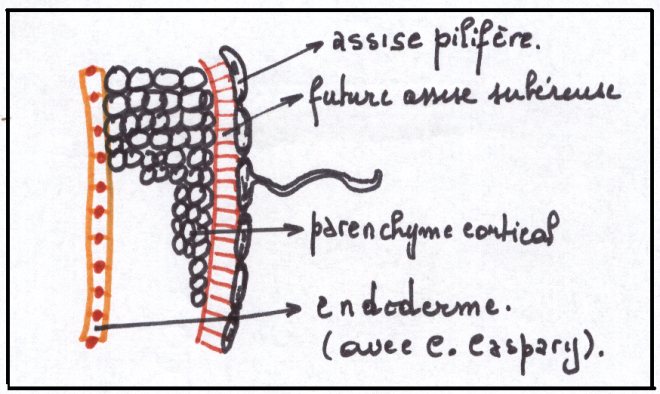
Entre l’épiderme et l’assise fibreuse existe du parenchyme externe .
Les assises de parenchyme externe les plus proches de l’épiderme sont
peu évoluées et capable de se rediviser , au besoin après avoir subi une
dédifférenciation, c’à d qu’elles redeviennent méristématiques .
C’est au niveau des cellules du parenchyme externe - cellules sous
épidermiques , mais parfois au niveau du parenchyme plus profond – que
la plante produit un néoméristème.
Les cellules issues de ce néoméristème seront différenciées en liège ( pas de méats ni dans l’épiderme ni dans le liège)
Origine du phellogène et son fonctionnementSoit une cellule du parenchyme dédifférencié. Elle prend d’abord une
cloison parallèle à l’épiderme vers l’intérieur : la cellule fille
produite vers l’intérieur
deviendra parenchymateuse et est appelée
phelloderme ( le phelloderme comporte parfois plusieurs assises). La
cellule mère fonctionne comme une
initiale : c’est le phellogène.
Toutes les autres division du phellogène seront extérieures à ce
dernier, soit orientées vers l’épiderme.Les cellules filles
successivement formées refouleront les premières formées et un nouveau
tissus, évoluant par voie centripète, se constitue, c’est le
liègeLes cellules de liège apparaissent en lignes radiales ; elles sont
imprégnées de subérine ; leur protoplaste disparaît et elles se
remplissent d’air
Les cellules de liège ont des contours anguleux et sont évidemment améatique.
Le périderme est le nom donné à l’ensemble liège + phelloderme+phellogène
La durée du phellogène varie selon les espèces :
-
Phellogène permanent : fonctionne pendant toute la vie de la plante ( hêtre, charme,…. Plantes à écorce lisse)
-
Phellogène annuel :
exemple la vigne , dont la tige s’appelle cep. Dans le cep, la plante a
recours à des assises de parenchyme cortical de plus en plus
profondes . Ce dernier peut s’épuiser jusqu’au liber .
-
Phellogène temporaire : l’exemple le plus typique est celui des platanes ; il existe également d’autres espèces, par exemple certains érables ; les
bouleaux, Merisiers…
Platanes : le phellogène se dsquamme , les écailles se détachent en plaques laissent un fond jaunâtre.
Bouleaux , merisiers : le phellogène se détache en copeaux tangentiels
Groseillers : lanières longitudinales
Chez beaucoup d’espèces : le phellogène se conserve sur le tronc et l’écorce se crevasse profondément
L’épaisseur du liège est de mince à épaisse et même très épaisse
Mince : hêtre , charme
Assez mince : bouleau, merisier
Epaisse : ormes et surtout vieux chênes, vieux peupliers
Très épaisse : chêne liège , le chêne liège donne des épaississement qui ,en 10 ans , peuvent atteindre 10 à 15 cm
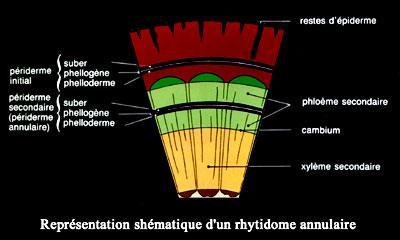
Le rhytidome est le nom donnéà l’ensemble des tissus morts extérieurs au phellogène , le phénomène est parfois assez complexe .
- Chez certaines plantes ( vigne) : chute annuelle ; bandes longitudinales
- Chez d’autres plantes ( érable, platane), élimination par plaques,
par copeaux longitudinaux( groseilliers) ou tangentiels ( bouleaux,
cerisiers)
- Chez beaucoup d’espèces, le rhytidome ne se desquamme pas et le
liège s’accumule .C’est dans ce cas que le phénomène est complexe.
En général, au fur et à mesure que le diamètre de la tige augmente,
de nouvelles zones génératrices de nouvelles zones génératrices
subéro-phellodermiques peuvent apparaître , cette fois – ci dans le
phloème secondaire . Elles produisent de nouvelles plaques de liège .
Les tissus situé à l’extérieur mourront et s’exfolieront , ce qui permet
au tronc de se développer.
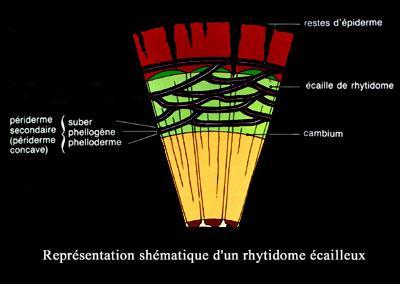
Avec un rhytidome écailleux, les écorces sont généralement plus
épaisses et se détachent du tronc sous formes d'écailles de tailles très
variables.
 L’écorce d’un arbre
L’écorce d’un arbre est un terme du langage courant qui, du point de vue anatomique s’adresse à l’ensemble des tissus extérieurs au bois.
Dans les vieux arbres ayant formé des assise subéro phellodermiques
en profondeur on distingue dans le rhytidome, de l’intérieur vers
l’extérieur les tissus
suivants : liber-phloème-fibres-phelloderme-phellogène-liège –( épiderme)
Dans cette succession, le rhytidome engloberait ( partiellement ou
non)les tissus allant de l’épiderme jusqu’au phloème et liber.
Lenticelles : le
manchon de liège ne peut être continu autour de la tige , il faut des
ouvertures pour permettre la respiration des cellules encore vivantes
à l’intérieur de la tige . Ces ouvertures sont des lenticelles constituées de cellules très turgescentes non subérisées.
Les lenticelles proviennent du phellogène qui à certains niveaux
donne des rangées de cellules filles qui deviendront parenchymateuses au
lieu de se subériser
Les lenticelles sont perpendiculaires à l’axe de la tige ou du rameau
et sont généralement situées à l’aplomb des anciens stomates dont elles
prolongent en
quelque sorte la fonction sans être aussi perfectionnée que ces derniers.
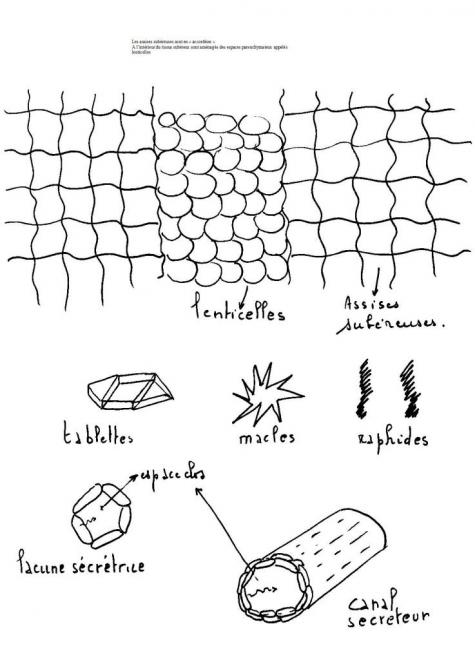
Le parenchyme le plus externe brunit, meurt et fait hernie par
l’ostiole qu’il déchire. Les lenticelles sont parfaitement visibles dans
un bouchon de liège.
On peut aisément reconnaître les lenticelles sur certaines tiges :
sureau, noisetier, sur certains troncs à écorce lisse sur laquelle la
lenticelle
s’étire transversalement ( ex : jeunes peupliers hybrydes euraméricains c. v serotina de champagne ).
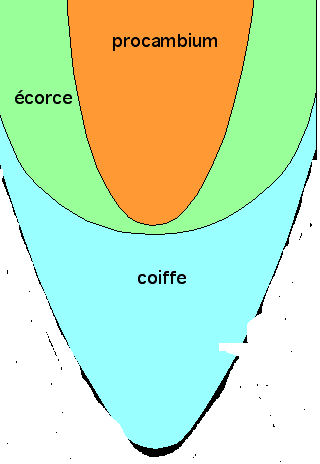
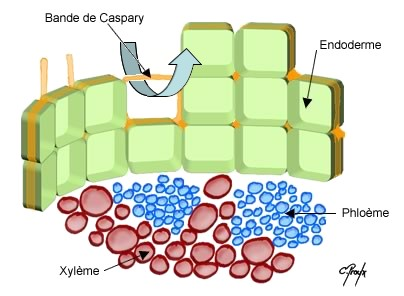
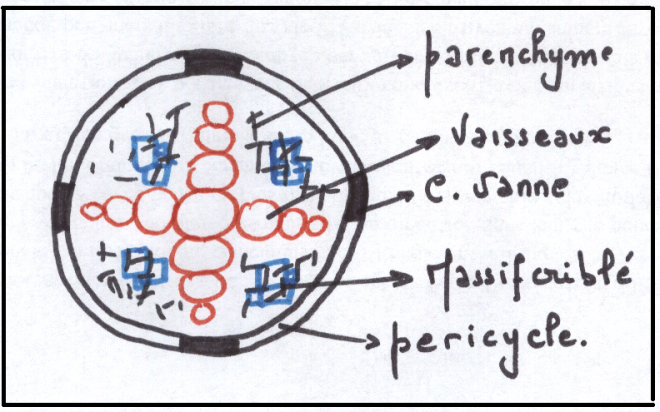
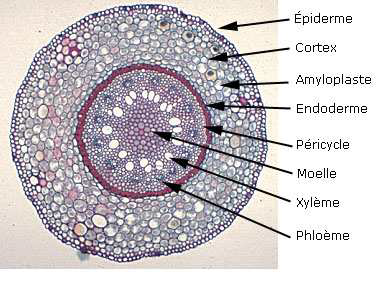
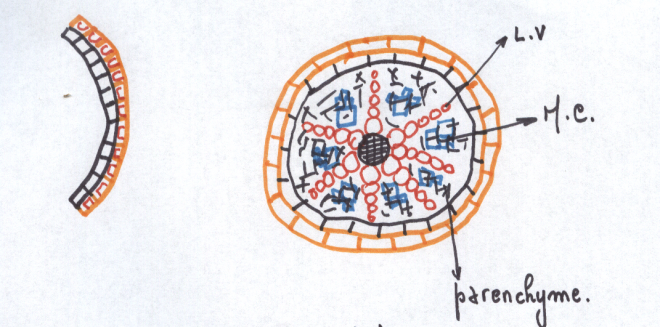
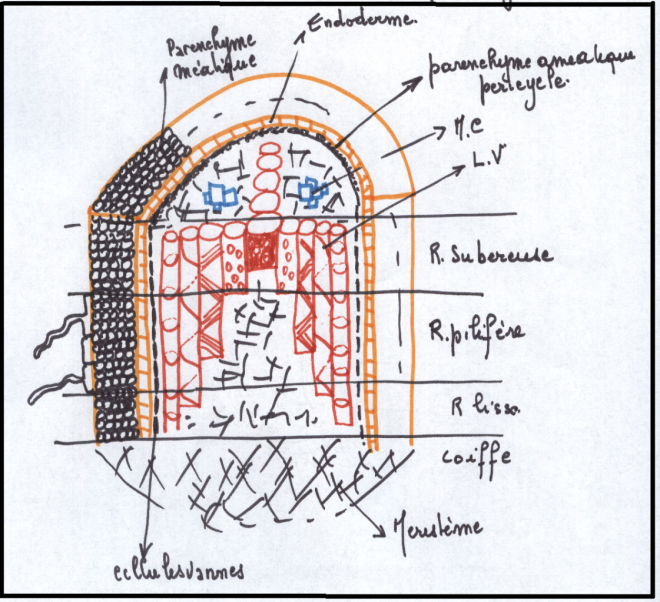
LA STRUCTURE SECONDAIRE DE LA RACINEIl faut évidemment que la racine s’épaississe corrélativement au
développement latéral de la tige . L’issue de ce développement est la
souche de l’arbre.
La structure secondaire se situe comme pour la tige à deux niveaux :
- Structure secondaire conductrice : production de bois et de liber
- Structure secondaire protectrice : production de liège
La conduction secondaire se réalise dans le cylindre central ; la
protection secondaire se réalise à l’intérieur du cylindre cortical.
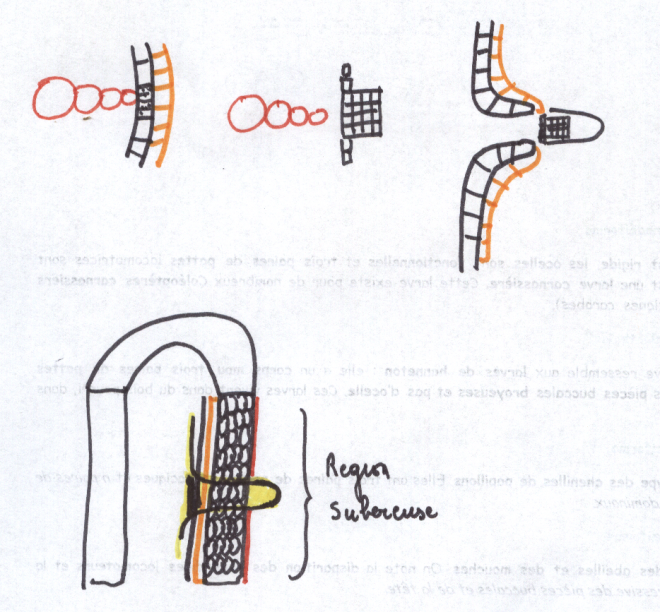
Réalisation d’un cylindre cambial radiculaire – son fonctionnementLe cambium fonctionne rapidement dans une tige susceptible d’accroissement secondaire .
Or dans une tige il n’existe pas de vaisseau donc pas de cambium
La racine doit donc réaliser un cambium afin de s’épaissir parallèlement à la tige
Les cellules améatiques du parenchyme central qui sous tend les
massifs criblés vers l’intérieur sont restée presque méristématiques .
Elles sont capable de se diviser à nouveau et de fonctionner comme les
initiales d’un méristème qui va se diviser à la manière du cambium.
Les cellules du cambium apparaissent d’abord comme des arcs cambiaux
autour des massifs criblés ; les arcs s’allongent s’allongent
latéralement vers le
péricycle, aux confins des lames vasculaires pôles ligneux.
Plus tard,au niveau des pôles ligneux, des cellules péricycliques
dédifférenciées construisent de nouveaux arcs cambiaux se raccordant aux
précédents. A ce
sujet, les versions varient quelque peu selon les
auteurs, en effet certains d’entre eux affirment que le péricycle ne
produit pas de cambium, mais que par
des simples divisions, il permet le raccordement. En tous les cas, c’est au niveau du péricycle que le raccord s’effectue.
Le circuit de cambium doit maintenant prendre sa forme circulaire.
On assiste à une inversion de courbure au niveau des massifs criblés
qui sont largement refoulés vers les tissus extérieurs, tandis que le
cambium fonctionne en produisant du bois et du liber , à l’instar de la
tige.
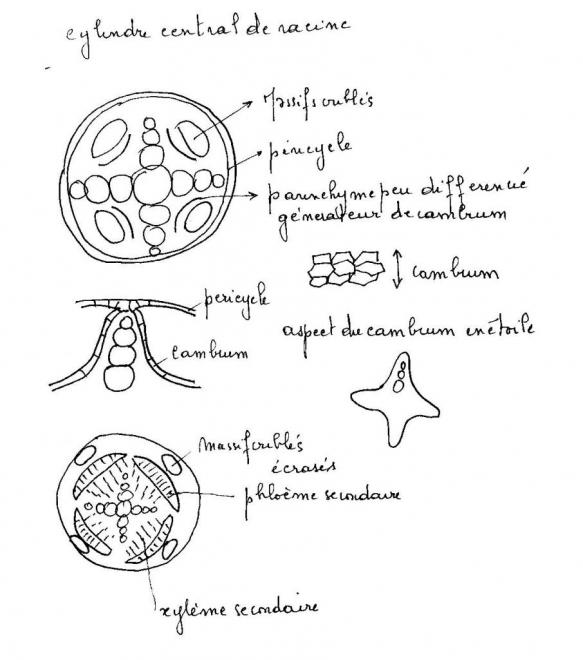 Réalisation d’un phellogène
Réalisation d’un phellogèneLes assises les plus externes du cylindre cortical sont généralement
peu méatiques. Elles réalisent un phellogène qui produira un liège, avec
lenticelles, comme dans la tige.





 2ème cas
2ème cas