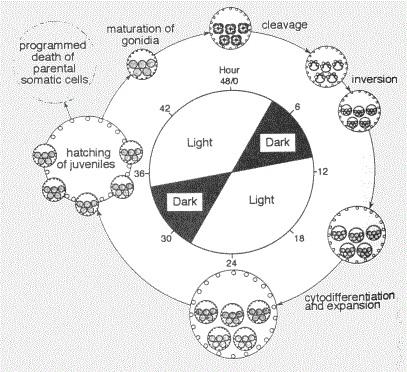
|
| | Classification.générale (cours) |  |
| | | Auteur | Message |
|---|
biologista

Sexe : 
 Messages : 180 Messages : 180
 Points : 317 Points : 317
 Date de naissance : 07/05/1992 Date de naissance : 07/05/1992
 Age : 32 Age : 32
 Emploi : Chez mon papa Emploi : Chez mon papa
| Sujet: Classification.générale (cours) 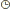 Mer 5 Déc 2012 - 23:21 Mer 5 Déc 2012 - 23:21 | |
| Classification générale
source: biodis
I.METHODES GENERALES DE CLASSEMENT DES PLANTES Il
existe différentes manières de classer les plantes , selon le point de
vue, le but, l'époque, les divers progrès de la science, les auteurs,
etc...On peut considérer, grossièrement, deux tendances générales :I.1 METHODE NATURELLEBasée sur la notion d'affinité. Elle réunit des plantes manifestement apparentées......selon l'évolution des critères. Une des plus anciennes classifications est celle de Antoine Laurent de Jussieu, botaniste de Louis XV.Actuellement,
on établit des classifictions qui s'efforcent de réunir des taxons
(noms) de plantes en les intégrant dans une sorte "d'arbre généalogique"
retraçant l'évolution végétale dans les temps géologiques, chaque
branche de l'arbre généalogique constituant un Phyllum. Les phyllums sont plus ou moins ramifiés ; les végétaux actuels sont évidemment situés aux extrémités des ramifications ...Ce mode de classification est issue d'une perspective phylogénétique :Voir l'ouvrage " Botanique systématique, une perspective phylogénétique" chez De Boeck Université I.2 CLASSEMENTS ARTIFICIELSC'est
la perspective adoptée par la plupart des flores simples, c'est à dire à
la portée de tous, quelle que soit la formation botanique de
l'utilisateur. Les plantes sont groupées en fonction d'un seul ou de
plusieurs caractères communs. Ces classements réunissent des plantes qui
n'ont aucunes affinités entre elles ; ils ont toutefois l'avantage
d'être pratiques. Ainsi, par exemple, LINNE a établit un classement des
phanérogames en tenant compte du nombre d'étamines et de carpelles. NB.
Il existe des flores à caractère sientifique qui les rapprochent des
classements naturels; leur utilisation est parfois ardue et postule des
bases botaniques réelles.I.2 LES ENBRANCHEMENTS  algues.pdf - Classiquement on distingue 4 voire 5 embranchements : algues.pdf - Classiquement on distingue 4 voire 5 embranchements :
- (Les Cyanophytes)
= organismes unicellulaires schyzophytes procaryotes proches des
Bactéries,ils sont chlorophylliens ( Algues bleues ), certains auteurs
ne les reprennent pas dans le monde végétal
- Les Thallophytes
- Les Briophytes
- Les Ptéridophytes,
ces trois groupes :Thallophytes, Briophytes, Ptéridophytes sont
regroupés en Cryptogames ( mode de reproduction cachée : spores)
- Les Spermatophytes, ce sont des organismes aussi appelés Phanérogames , à mode de reproduction "apparente" : graines.
On distingue également les Thallophytes par opposition aux Archégoniates : Les Thallophytes,
dont les Algues et les champignons. L'appareil végétatif est un
Thalle ; la paroi protectrice des gamétanges femelle est une membrane. Les Archégoniates
: l'appareil végétatif est composé, s'il est complet, de tiges,
feuilles et racines; le gamète femelle est entouré par une assise
cellulaire en forme de bouteille appelée Archégone.
A noter qu'actuellement le terme "Cormophytes" ( plantes possédant un
"cormus") correspond aux mêmes organismes que le terme "Archégoniates",
puisque l'on considère que certains Briophytes et les Ptéridophytes
possèdent à un moment de leur cycle vital un rudiment de cormus ou un
cormus plus ou moins complet. Anciennement, le terme "Cormophytes"
correspondait au terme "Phanérogames". Le cormus est un appareil
végétatif comportant tige feuilles et racine. 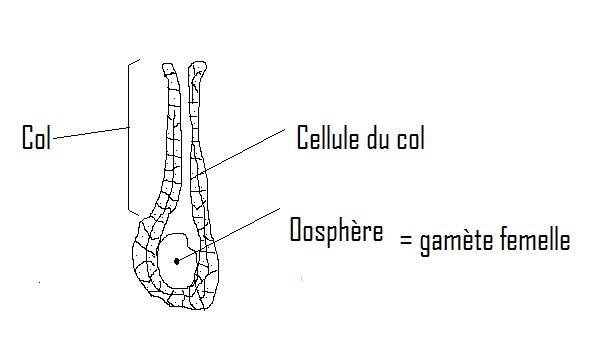 Schéma : Archégone.Les Archégoniates se décomposent composent en :- Briophytes, Schéma : Archégone.Les Archégoniates se décomposent composent en :- Briophytes,
dont les mousses : plantes collées au sol, sans racines ni tissus
conducteur; à cycle haplo-diplobionte dont la phase haploïde est
dominante.- Ptéridophytes,
dont les Fougères : ayant un tissus conducteur ( = Cryptogames
vasculaires ); à cycle haplo-diplobionte dont la phase diploïde est
dominante.- Spermatophytes, plante ayant, en plus des caractères précédents des graines ( et des fleurs chez les Angiospermes ). On
doit noter différentes tendances actuelles de classer en distinguant
plusieurs règnes : Le Professeur PIERRART a été à l'origine de la
distinction du règne des procaryotes ( bactéries diverses ); des
protistes ( protozoaires , phycomycètes, algues) ; des végétaux ( algues
exclues ); des champignons non flagellés et des animaux. Une autre
classification est celle de WITTHAKER en cinq règnes : animal, végétal (assemblage
polyphylétique d’organismes photosynthétiques et dont la paroi est
faite de cellulose. Ce groupe est formé de deux lignées, l’une d’algues,
et la seconde de plantes terrestres, qui comprennent notamment les
bryophytes (mousses et hépatiques), fougères (ptéridophytes),
gymnospermes et angiospermes), champignon(les
champignons vrais appartiennent au groupe des Eumycètes et se
répartissent au sein des Basidiomycètes, comme les amanites ou, pour
quelques-uns, parmi les Ascomycètes, à l'exemple des morilles ou des
truffes. L'étude de ces champignons est la mycologie et ceux qui la
pratiquent sont des mycologues. Mais il y a aussi d'autres groupes qu'on
appelle communément champignon mais qui n'en sont pas au sens strict du
terme qui sont les oomycètes (plus proches génétiquement des algues
brunes) et les myxomycètes), protiste (réunit
des organismes à organisation cellulaire dite simple, unicellulaires le
plus souvent, multicellulaires parfois mais sans tissus spécialisés.
Certains sont autotrophes (p. ex. : microalgues), d'autres sont
hétérotrophes (p. ex. : protozoaires se nourrissant de microalgues).
Les protistes sont un groupe paraphylétique dans la classification
phylogénétique. Ils étaient, en classification classique, le quatrième
règne du domaine des eucaryotes.) et monère (Procaryotes) .Il va de soi que la notion classique d'Embranchement est alors bouleversée dans son niveau classique.Exemple : II. L'EMBRANCHEMENT DES THALLOPHYTES II.1 GENERALITES Traditionnellement II. L'EMBRANCHEMENT DES THALLOPHYTES II.1 GENERALITES Traditionnellement
l'on considère que les Thallophytes sont des organismes à caractères
morphologiques négatifs : pas de racines, pas de tiges pas de feuilles
pas de graines. La tendance actuelle est d' admettre l'existence chez
les thallophytes de structures proches de tiges mais sans système
conducteur ( ça équivaut à un thalle) parfois parfois le terme tige est
utilisé.De manière générale nous dirons donc que l'appareil végétatifde ces plantes est constitué par un THALLE;Caractères généraux du thalle . Très variables à savoir :Forme
: du bâtonnet ou de la sphérule microscopique ( bactérie, les bactéries
sont parfois classées dans les thallophytes )jusqu'à une forme rubanée
longue de 200 à 300 mètres ( certaines algues brunes ). Structure : - Cellulaire- articulaire- symplastique Organisation : uni ou pluricellulaire de différents types :- filamenteux- lamellaire- massif- coloniaire Organisation : uni ou pluricellulaire de différents types :- filamenteux- lamellaire- massif- coloniaire Noyau : parfait ou primitifMembrane cellulaire :- plasmatique: membrane zone périphérique de la cellule plus dense- cellulosiqueMode de reproduction :- multiplication végétative- reproduction agame ( spores)- reproduction sexuéeCycles de développement :- diplophasique- haplophasique- haplo-diplophasique Physiologie - organismes autotrophes : pourvus de chlorophylle ( exemple : algues ).- organismes hétérotrophes : pas de chlorophylle ( ex: champignons ).Ces derniers sont parasites, saprophytes ou symbiotiques Conclusion générale : Embranchement très hétérogène que certains auteurs n'ont pas hésité à fragmenter.Points communs aux Thallophytes ( ou caractères positifs)1.Les organes de reproduction sont enfermés dans une membrane, la membrane étant étant opposée à la paroi assisiale des archégoniates.2.La division cellulaire est réalisée par voie centripète ( exemple:scissiparité) tandis qu'elle est centrifuge dans les Embranchements supérieurs .Rm: elle est également centripète chez les animaux. II.2 LE GROUPE DES THALLOPHYTES 1. Algues : La Noyau : parfait ou primitifMembrane cellulaire :- plasmatique: membrane zone périphérique de la cellule plus dense- cellulosiqueMode de reproduction :- multiplication végétative- reproduction agame ( spores)- reproduction sexuéeCycles de développement :- diplophasique- haplophasique- haplo-diplophasique Physiologie - organismes autotrophes : pourvus de chlorophylle ( exemple : algues ).- organismes hétérotrophes : pas de chlorophylle ( ex: champignons ).Ces derniers sont parasites, saprophytes ou symbiotiques Conclusion générale : Embranchement très hétérogène que certains auteurs n'ont pas hésité à fragmenter.Points communs aux Thallophytes ( ou caractères positifs)1.Les organes de reproduction sont enfermés dans une membrane, la membrane étant étant opposée à la paroi assisiale des archégoniates.2.La division cellulaire est réalisée par voie centripète ( exemple:scissiparité) tandis qu'elle est centrifuge dans les Embranchements supérieurs .Rm: elle est également centripète chez les animaux. II.2 LE GROUPE DES THALLOPHYTES 1. Algues : La
plupart sont aquatiques ( marines ou d'eau douce ) , àn l'exception de
quelques algues vertes ou bleues vivant sur les rochers , écorces
d'arbres, pierres humides, toitures etc...Toutes les algues possèdent de la chlorophylle ( parfois masquée par un autre pigment ) : elles sont donc autotrophes .2. Champignons : sont dépourvues de chlorophylle = plantes hétérotrophes .- Parasites : vivent sur des tissus végétaux ou animaux vivants.- Saprophytes
: vivent au dépens de la matière organique morte, et contribuent à
minéraliser la matière organique dont ils débarassent la surface du sol.- Symbiotiques : association à bénéfice réciproque entre une plante et un champignon.3. Lichens : Plantes aériennes : association d'une algue et d'un champignonREMARQUES-
Les microbes sont des Protozoaires s'ils sont classés dans le règne
animal; dans le règne végétal ils se répartissent en champignons,
bactéries ou virus.- Les virus
ne sont pas examinés dans cette rubrique, nous signalons toutefois
qu'ils sont des corps chimiques inertes très petits ( inférieurs à 0,001
mm ) qui ne s'animent et se multiplient qu'au contact d'une cellule
vivante ( plante, animal, bactérie ). Ils sont responsables de maladies
de leurs cellules hôte ou organismes hôte et sont souvent véhiculés par
les pucerons ( agronomie ). II.1.LES ALGUESElles sont réparties en plusieurs classes qui marquent leur degré d'évolution . CLASSE I : LES ALGUES BLEUES OU CYANOPHYCEES - CYANOPHYTA , voir la rubriques "algues"Primitives, pas de noyau parfait et la chlorophylle n'est pas fixée sur un stroma : elle est diffuse dans le cytoplasme.Algues pourvues d'un pigment
bleu : la phycocyanine, associée à la chlorophylle, associé à la
chlorophylle ( parfois aussi un pigment rouge, la phycoérithrine mais
sont associées aux algues bleues en raison de leurs caractères primitifs
). 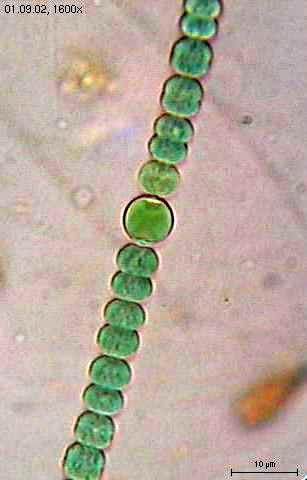 Reproduction Reproduction
végétative par scissiparité si elles sont unicellulaires, sinon c'est
une des cellules de la colonie ou du filament qui se fractionne pour son
propre compte.Habitat: eaux douces, sols humides, rochers humides.Exemple : Nostocs ou crachats de lune .
Intérêts.
Les algues bleues sont capables de fixer l'azote atmosphérique, tout
comme certaines bactéries ( nodosités fixatrices d'azote chez les
plantes de l'ordre des légumineuses ) ; surtout importantes dans les
rizières. CLASSES II : LES FLAGELLATES = EUGLENOPHYTATrès simples, parmi les plus inférieures, mais elles ont un noyau parfait avec des chloroplastes. Elles sont plus évoluées que les précédentes 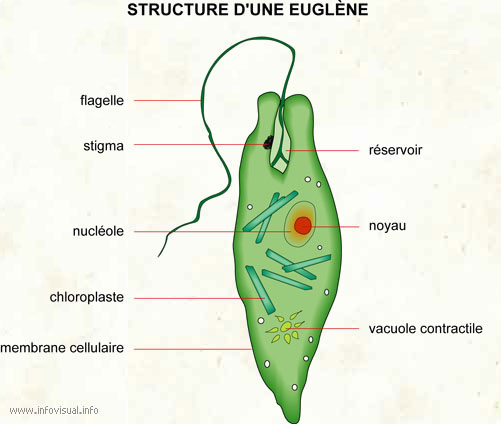 Le genre le plus typique est Le genre le plus typique est
l’Euglena, courant dans les étangs et les piscines, spécialement
lorsque leurs eaux sont polluées par des engrais provenant de leur
environnement.Lesflagellésde
ce groupe, qui comprend environ mille espèces, se trouvent couramment
dans les eaux douces chargées de matières organiques ; il existe
également quelques espèces marines etendosymbiotiques.La reproduction se fait par division cellulaire longitudinale.Ce
sont des organismes qui sont aussi à classer parmi les protozoaires
chlorophylliens, raison pour laquelle nous les étudions içi. CLASSE III. LES DIATOMEES = ALGUES SILICEUSES = BACILLARIOPHYTAAlgues
microscopiques et toujours unicellulaires, rencontrées surtout en eaux
marines, mais également en eaux douces et sur la terre humide. Enfermées
dans une sorte de carapaces à deux valves ( qui rappelle l'aspect d'une
boîte de Pétri ) et de forme variable, avec des dessins en relief
silicifié, constants pour chaque espèce.Reproduction sexuée, par isogamie ; cycle diplobionte ( = diplophasique ). Les valves Les valves
sont composées de substances pectiques imprégnées par de la silice qui
les rend très dures et inextensibles; elles persistent après la mort
des organismes. Leur accumulation constitue des sédiments géologiques
qui sont utilisés ( plus par le passé) pour différents emplois :
polissage ders métaux ( abrasifs), filtration, isolants.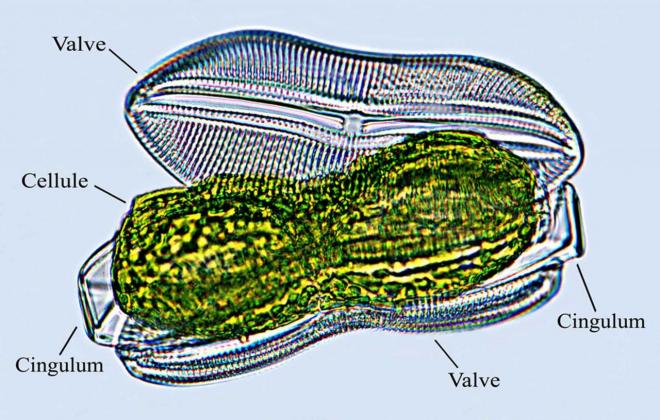 Composition : - deux valves - cytoplasme + noyau parfait- deux grandes plaques brunâtres = chloroplastes ( mais pas classées dans les algues brunes) masqués par la fucoxanthine. - goutelettes d'huile dans le cytoplasme.Ces animaux sont doués de mouvements dont les mécanismes ne sont pas encore parfaitement élucidés. Composition : - deux valves - cytoplasme + noyau parfait- deux grandes plaques brunâtres = chloroplastes ( mais pas classées dans les algues brunes) masqués par la fucoxanthine. - goutelettes d'huile dans le cytoplasme.Ces animaux sont doués de mouvements dont les mécanismes ne sont pas encore parfaitement élucidés. 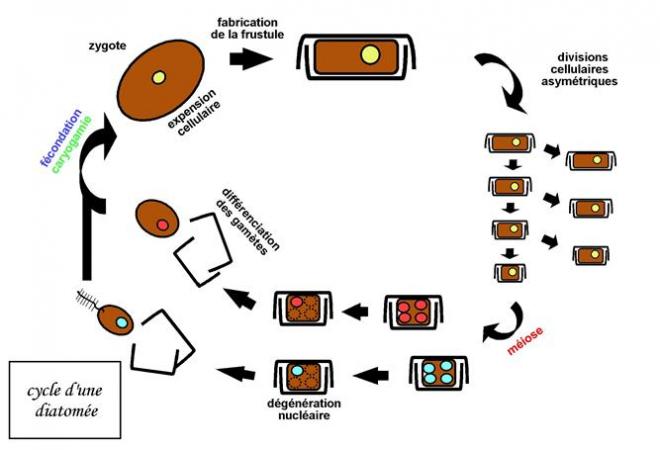 CLASSE IV. LES ALGUES VERTES OU CHLOROPHYCEES C'est un groupement d'organismes très hétérogène :1. Variété des thalles-unicellulairesSpirogyre : La Spirogyre est CLASSE IV. LES ALGUES VERTES OU CHLOROPHYCEES C'est un groupement d'organismes très hétérogène :1. Variété des thalles-unicellulairesSpirogyre : La Spirogyre est
une algue verte filamenteuse commune dans les fossés et les mares d'eau
suffisamment pure. Longue de plusieurs décimètres, elle est formée par
un filament non ramifié de 50 à 100 mm de largeur, fait d'un
enchaînement linéaire de cellules rectangulaires pourvues chacune d'un
ou de plusieurs chloroplastesen
forme de ruban spiralé. Ces longs filaments ondulent très lentement
dans l'eau, ce qui permet le rapprochement des individus, nécessaire à
la reproduction sexuée. hétérogamie, hétérothalleChlamydomonas-pluricellulairesVolvox: oogamieUlothrix: isogamie-coloniairesPediastrum: agameSenedesmus: isogamie-articulairesCladophora : seul haplodiplobionte.-symplastiquesVaucheria 2. Reproduction Sexuée ou à la fois agame et sexuée ( avec ou sans alternance de l'un ou l'autre mode de reproduction). 3. Mouvement : douées de mouvements ( flagelle ) ou non 4. Fécondation
: par oogamie, isogamie, ou hétérogamie, ces derniers cas sont des
plantes homothalle ( un individu porte les deux sexes ) ou hétérothalle (
un sexe par individu ). 5. Quelques cycles biologiques de Chlorophycées haplobiontes ( haplophasique). Spirogyre : hétérothallie, hétérogamie 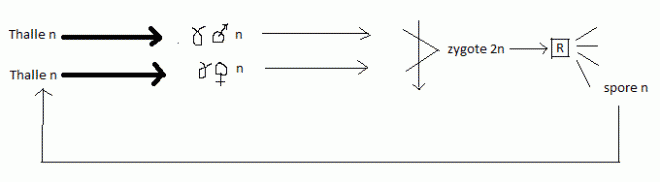 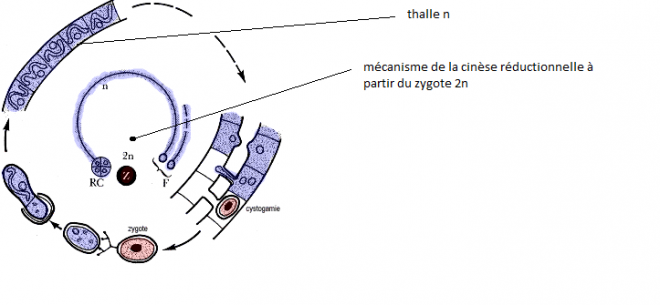 D'après Lamotte M. L'Héritier P.. 1965. Biologie générale. Doin, Paris, t1 : 100.De manière plus schématique D'après Lamotte M. L'Héritier P.. 1965. Biologie générale. Doin, Paris, t1 : 100.De manière plus schématique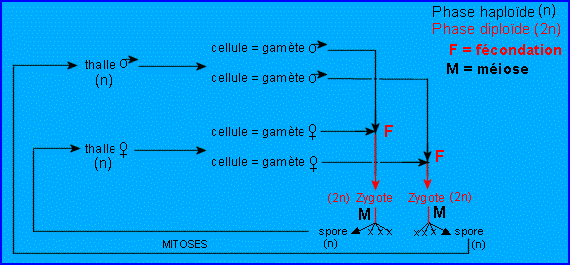 Chlamydomonas: isogamie homothalle Chlamydomonas: isogamie homothalle 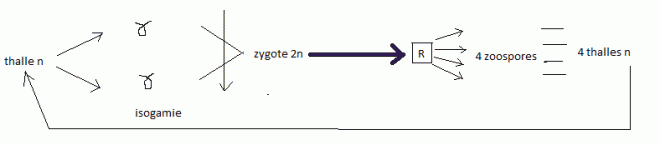 Ces algues unicellulaireminuscules (d'une taille de10 µmenviron) sont munies de deux flagelles, et d'unchloroplasteunique en forme de cloche lui permettant d'opérer la photosynthèse.C'est un des genres retenus pour la production éventuelle debiocarburants(hydrogène ou hydrocarbure) par des algues. Ces algues unicellulaireminuscules (d'une taille de10 µmenviron) sont munies de deux flagelles, et d'unchloroplasteunique en forme de cloche lui permettant d'opérer la photosynthèse.C'est un des genres retenus pour la production éventuelle debiocarburants(hydrogène ou hydrocarbure) par des algues.
Depuis les années 1960, l'espèceChlamydomonas reinhardtiiest
unorganisme modèle, de plus en plus utilisé en recherche fondamentale
et appliquée, notamment pour étudier les gènes et mécanismes
moléculaires impliqués dans laphotosynthèse, la nutrition ou encore la
motilité des flagelles.  La La
cellule a enkystée représente une forme de résistance qui permet à
l'espèce de traverser les périodes défavorables à l'état de vie
ralentie. Lorsque les conditions sont favorables, son enveloppe se rompt
et laisse échapper quatre cellules qui grandiront pour devenir
semblables à la cellule b.La
cellule b donne naissance à un clone par divisions successives. Dans
certaines conditions, une cellule b donne naissance à huit cellules c
plus petites que la cellule initiale mais présentant la même
morphologie.Deux cellules c issues de clones différents s'accolent puis fusionnent pour donner la cellule a.Ulothrix : homothallie, isogamie 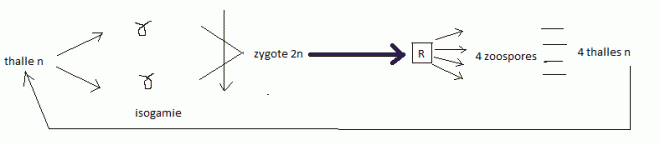 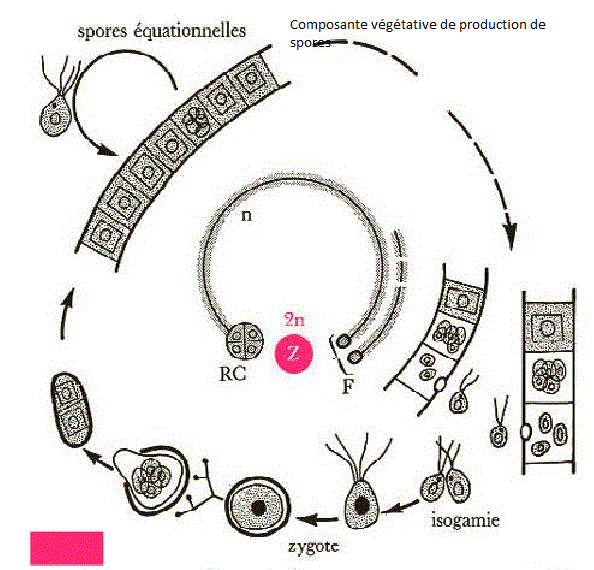 Volvox : homothallie oogamie ( reprouction asexuée et sexuée ) Volvox : homothallie oogamie ( reprouction asexuée et sexuée )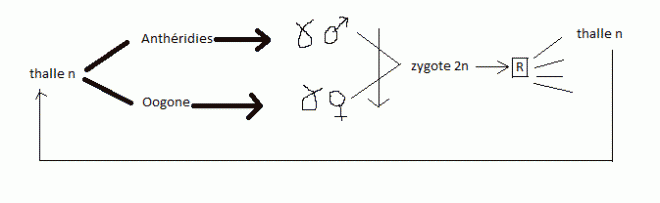 Chaque
colonie est constituée en fonction des espèces de 500 à 60 000 cellules
biflagellées qui ressemblent à chlamydomonas (les mouvements
synchronisés des flagelles permettent le mouvement de l'ensemble de la
colonie, cf films 1et 2). et de 16 cellules germinales (chez V. carteri).
La différentiation en cellules somatiques et germinales est liée à une
division cellulaire asymétriques (la grosse cellule devenant la cellule
germinale). Il semble que la taille cellulaire soit le déterminant
principal des différentiations et des études moléculaires sont menées
actuellement pour déterminer les mécanismes moléculaires mis en jeu. Les
cellules germinales gagnent l’intérieur de la colonie et des
mini-colonies se développent à l'intérieur de la colonie principale. Les
mini-colonies ont les flagelles tournés vers l'intérieur. Il se produit
un retournement des mini-colonies (comme le retournement d'un doigt de
gant) pour orienter correctement les flagelles. Puis, la colonie mère
soit meurt, soit se perfore libérant les nouvelles colonies. Cette algue
peut aussi faire la reproduction sexuée, pendant laquelle quelques
cellules se différencient en "ovules" de grande taille et d'autres en
"spermatozoïdes" de plus petite taille. Du fait de sa structure
coloniale, Volvoxsert de modèle d'études pour les premières étapes menant à la pluricellularité. Vaucherie : homothallie, oogamie ( reproduction asexuée et sexuée ) 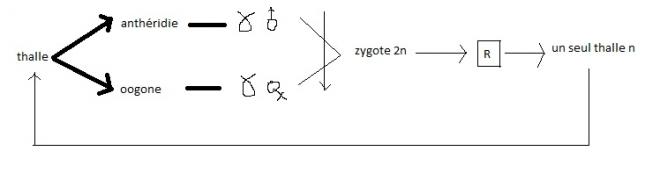 | |
| | | | biologista
Sexe :
Messages : 180
Points : 317
Date de naissance : 07/05/1992
Age : 32
Emploi : Chez mon papa
| | Sujet: Re: Classification.générale (cours) Mer 5 Déc 2012 - 23:29 | |
| * Rappel vocabulaireisogamie
: Mode de reproduction sexué dans lequel les deux gamètes sont
identiques, de telle façon qu'on ne puisse distinguer le mâle de la
femelle.L'isogamie parfaite se rencontre généralement chez les champignons.anisogamie : Se dit quand un gamète (mâle ou femelle) est plus gros ou différent.oogamie
: Mode de fécondation de nombreuses espèces animales et végétales,
consistant en un unique processus d'union entre deux cellules
différentes, un gamète femelle, oosphère immobile et un gamète mâle,
spermatozoïde mobile flagellé et comprenant surtout un noyau.II.2. LES CHAMPIGNONS ou Mycètes ou FungiL'étude des
champignons couvre à elle seule un chapitre important de la
classification des plantes. Elle fait l'objet d'une discipline
particulière :II.2.1 LA MYCOLOGIEGénéralités 1. Beaucoup de Champignons ( et d'Algues ) présentent une structure filamenteuse parce
que les cellules sont allongées et les divisions successives sont
orientées de manière à ce que les cellules se succèdent bout à bout.2. Les filaments ou hyphes sont plus ou moins imbriqués les uns dans les autres ; ils constituent, par leur réunion, non pas un tissus, mais un mycélium. 3. Dépourvus de chlorophylle, les champignons sont hétérotrophes et soit :- Parasites : responsables de nombreuses maladies des plantes et de quelques maladies des animaux ( "muguet", pelade ...)- Saprophytes : ils "minéralisent" la matière organique. Voir pédologie.-Symbiotiques : ils vivent en association avec d'autres plantes ( par exemple : mycorhises ). 4. la membrane de nombreux champignons contient de la chitine ; voir les animaux Arthropodes . 5. La substance
de réserve des champignons n'est pas constituée par de l'amidon, mais
par du glycogène ou bien des glucides simples ( sucres ). Le glycogène
est la forme de réserve des animaux. 6. La
reproduction se fait par spores , parfois zoospores, après un processus
sexué; ou par conidies ( reproduction asexuée : extrémité des hyphes se
ramifiant en cellules plus ou moins renflées sphériques.7. Estimation numérique : +/- 50000 espèces mycologiques : voir ci-après ( certains avancent de 70.000 à 90.000 espèces )8. Importance agronomique : a) Parasite, saprophytes ou symbiotiquesb) Industrie des levures et fermentations ( brasseries, distilleries, boulangeries ...c) Productions diverses, : antibiotiques ( ex : pénicilline, ), fromages raoquefort),...d) Alimentation humaine, champignons comestibles .Mais il y a aussi des champignons indigestes et surtout des champignons mortels.9. Les champignons inférieurs sont souvent étudiés en microbiologie .10. On peut classer les champignons comme suit : - Champignons non authentiques I. Bactéries ou schyzomycètes II. Souvent repris ailleurs, les Myxomycètes ( Archimycètes ) +/- 500 Espèces .- Champignons authentiques ou EumycètesIII. Phycomycètes ( Syphomycètes, Oomycètes ou encore Zygomycètes importants en phytopathologie )IV. AscomycètesV. Basidiomycètes - Fungi imperfecti (Champignons non définis par leur mode de reproduction sexuée)Encore appelés Adélomycètes, Hyphomycètes ou Deutéromycètes.Les
schyzobactéries ou schyzophytes comprennent les Cyanobactéries ou
Cyanophytes ( algues bleues ) mais aussi les organismes appelés
schyzomycètes, parfois repris avec les Bactéries .CHAMPIGNONS NON AUTHENTIQUESII.2.2 CLASSE DES SCHYZOMYCETES à caractères bactériens - Organismes microscopiques, unicellulaires. - Habitat : la biosphère, eau - sol - atmosphère.- Nombre d'Espèce relativement réduit par rapport aux Eumycètes. -Thalle de forme
variable : bacilles, coques ( Streptocoques, Staphylocoques...),
vibrions, spirilles... , parfois ciliés ou flagellés- Noyau : primitif ( diffus dans le cytoplasme ).- Membrane cytoplasmique souvent entourée d'une gaine gélatineuse : possibilité d'accolement. - Reproduction par bipartition : divisions extrêmement rapides si les conditions leurs sont favorables. - Si les conditions sont défavorables, enkystement ( gélatine), ce qui leur donne une résistance extraordinaire performances de
certaines bactéries : jusque 110°C pendant trente seconde en milieu
humide, jusque 180°C pendant trente secondes en milieu sec, jusque
-250°C on retrouve des bactéries vivantes , en général le froid ne tue
pas, il " endort " , ne pas confondre avec les actions bactériostatiques
et bactéricides .La
classification des bactéries, repose sur l'examen de différent critères (
milieux de culture ) comme le potentiel de dégradation de certaines
molécules, le caractère aérobie ou anaérobie, critères biochimiques,
mais aussi les caractères parasites, saprophytes ...II.2.3 CLASSE DES MYXOMYCETES OU ARCHIMYCETES Champignons inférieurs, parfois "disputés" entre zoologistes et botanistes.Par exemple Fuligo septica : Myxomycète saprophyte sur vieux cuirs.Cycle résumé :Une spore
( haploïde ) germe en libére son cytoplasme et devient une zoospore.
La zoospore prend une forme qui rapelle celle d'une Amibe avec
pseudopodes ( nutrition et déplacement ). Eventuellement elle peut
s'enkyster en conditions défavorables. Au moment donné , le noyau (n) se divise et les gamètes fils obtenus sont des gamètes (n).Les gamètes ( homothallie ou hétérothallie selon les espèces ) se réunissent pour donner un noyau 2n ou zygote Les zygotes se
réunissent pour donner un gros symplaste ( appelé plasmode ) doué de
mouvements de reptation. Ce symplaste est en réalité un sporange dont
les noyaux 2n subissent des cinèses réductionnelles et libèrent des
spores ( haploïdes).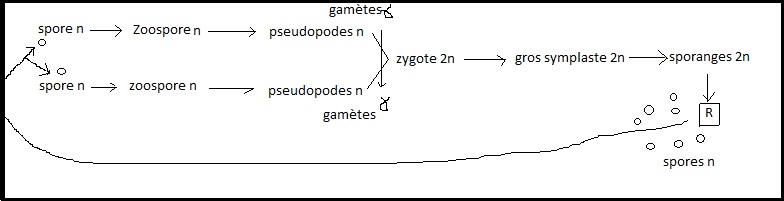 (Dans ce cycle précis il y a hétérothallie) Remarque (Dans ce cycle précis il y a hétérothallie) Remarque
: Certains auteurs classent parfois Plasmodiophora brassicae (ordre
des Plasmodiophorales ) parmi les Myxomycètes. D'autres les situent dans
les Phycomycètes : le classement semble difficile, Plasmodiophora
brassicae est l'agent responsable de la hernie du chou,
qui attaquent en réalité les racines de nombreuses plantes de la
famille des Brassicacées ( anc. Crucifères ) où elle peut causer des
dégâts extrêmement dommageables aux cultures maraîchères ( Choux ),
agricoles ( Navets ), ou ornementales ( Giroflées, Arabis, etc...).Autres
Myxomycètes : Synchytrium endobioticum ( agent de la gale noire ou
maladie verruqueuse de la pomme de terre ); Eocarpus fragilis ; etc...CHAMPIGNONS VRAIS OU EUMYCETES Appareil végétatif filamenteuse : feutrage de filaments appelé hyphes qui sont réunis en un mycélium .A certains moments, l'on constate que certains filaments se redressent et se rassemblent en un corps massif ou stroma , qui est le siège de la reproduction, à tout le moins pour les deux classes supérieures.- classes des Phycomycètes: hyphes symplastiques ( cytoplasme indivis )- classe des Ascomycètes : hyphes symplastiques ( = 1 noyau par cytoplasme )- classe des Basidiomycètes : hyphes articulaires ( = plusieurs noyaux dans un même cytoplasme ) II.2.4 CLASSE DES PHYCOMYCETES OU SIPHOMYCETESChampignons
microscopiques à hyphes symplastiques souvent appelés "moisissures". Ces
champignons font penser à des algues ( d'où leur nom... ); ils ne
forment jamais de thalle massif comme les Asco. ou les Basidiomycètes.On y rencontre surtout des saprophytes, parfois des parasites des plantes vertes ou des animaux. Classification - Sous-classe des Oomycétales : reproduction sexuée par oogamie ( petit gamète mâle flagellé, gamête femelle plus volumineux ) ou ChitridiomycètesOrdres : Plasmodiophorales - Saprolégnales - Pérénosporales.- Sous-classe des Zygomycétales : reproduction sexuée par isogamie (en botanique, l'isogamie correspond aux végétaux gamètes mâles et femelles, flagellés, mobiles et de taille identique). ou Zygomycètes Ordres : Mucorales - Entomophtorales Plasmodiophorales : P.brassicae provoque la " Hernie du chou " . Déjà cité .Saprolégnales :
champignons vivant dans l'eau ; saprophytes ( sur certains déchets
organiques ) ou parasites ( sur certains poissons cyprinidés [carpes,
tanches ] ) S. parasiticaprovoquant la maladie appelée " la mousse " ou
"le blanc " zoospores biflagellés. Pérénosporales : - Plasmopora viticola
: champignon responsable du " Mildiou de la vigne " - Mildiou de "
meal-dew - : c'est le plus grand ravageur de la vigne ( feuilles et
raisins ) après le Phylloxera ( puceron ) . Il a été combattu avec des
bouillies à base de cuivre dont la célèbre " bouillie bordelaise " ,
mélange de Ca(OH)2 et CuSO4-----> CaSO4 ( gypse) + Cu(OH)2 .
- Phytium de Baryanum : est un des agents de la "fonte des semis" et
des boutures d'espèces très diverses : disparition des jeunes plantules
aussitôt après la germination. - Phytophtora infestans
: cause le "Mildiou de la pomme de terre et de beaucoup de Solanacées.
On en connaît qu'une reproduction végétative : il y aurait perte de la
reproduction sexuée. Le champignons persiste d'une année à l'autre dans
les tubercules de pommes de terre , ion le combat actuellement par des
fongicides dits systémiques à appliquer à plusieurs reprises en cours de
végétation, surtout en années pluvieuses .Mucorales : - Mucor mucedo
: moisissure apparaissant sur du pain humide, des vieilles confitures,
etc... . On lui reconnaît un mode de reproduction agame et un mode de
reproduction sexuée.a) Reproduction
agame : certains filaments e renflent et s'érigent, formant un sommet
en massue qui s'isole par une membrane transversale cellulosique. Une
tête apparaît, formant la columelle. Des spores se forment dans la
"tête" , sont libérées et forment de nouveaux filaments. De manière plus précise : cycle
haplobiontique agame . Les sporangiospores plurinucléées et haploïdes
germent pour donner naissance au mycélium. Celui-ci est peu cloisonné et
possède différents types d'hyphes: des hyphes horizontaux qui poussent
rapidement à la surface du substrat et qui sont assez massifs (les
stolons). Quand ils touchent la surface du milieu de culture, ils
donnent naissance à des hyphes plus grêles qui pénètrent le milieu et
assure la nutrition. Chez certaines espèces, la croissance pendant ce
stade en absence d'oxygène ou en présence de grande quantité de CO2
entraîne la germination des sporangiospores en forme levure. Le
mycélium différencie des sporangiophores qui sont des hyphes porteurs de
sporanges ( columelle) contenant eux-même les spores. Ces spores sont
plurinucléées et peuvent contenir jusqu'à 20 noyaux. Enfin, certaines
espèces différencient des spores de résistance à partir d'hyphes : les
chlamydospores.  b) Reproduction b) Reproduction
sexuée. A partir d'une spore (n) : hyphes mycéliens qui vont à la
rencontre l'un de l'autre ) forment chacun une " tête " qui s'isole ;
les "têtes" fusionnent ( structures de rencontre = suspenseurs ou
zygophores ). Tous les noyaux fusionnent deux par deux et donne un grand
zygote symplastique ( zygospore ) qui s'isole en s'entourant de
gélatine.ce zygote peut
donner soit un nouveau filament symplastique et les premières cinèses
sont réductionnelles ou alors donner un sporange dans lequel les premières cinèses sont réductionnelles et chaque spore donnera un filament . Plus précisément pour l'alternative du sporange : Chez M. mucedo,
il existe deux types sexuels (si la plupart des espèces de mucorales
sont hétérothalliques, quelques-unes sont homothaliques). En fait chacun
des types sexuels possède une voie incomplète de biosynthèse d'une
hormone, l'acide trisporique. Celle-ci est commune à toutes les
mucorales. Elle n'est fabriquée que lorsque les deux mycéliums mat+ et
mat- sont en présence. Lors de la rencontre de souches de types sexuels
compatibles, les mycéliums différencient sous le contrôle de l'acide
trisporique des structures particulières : les zygophores ou suspenseur.
Les zygophores plurinucléés croissent l'un vers l'autre et après un
processus de différenciation complexe fusionnent pour donner naissance à
une zygospore. Dans cette zygospore, les noyaux s'apparient par couples
puis fusionnent et les noyaux diploïdes obtenus font la méiose. Le
devenir des différents noyaux n'est pas toujours clair car en fonction
des espèces une zygospore donne naissance à des descendants d'un seul
type sexuel (cas de M. mucedo) ou à des descendants de l'un ou
l'autre type. La zygospore germe difficilement (de l'ordre de 1% de
germination) pour donner directement un sporangiophore qui va redonner
des sporanges. Celle-ci est particulièrement résistante et le faible
pourcentage de germination est probablement lié à un mécanisme de
dormance levée progressivement au cours du temps.C'est une plante haplobionte ( cycle haplophasique ) , Mucor mucedo est homothallique. Entomophtorales içi repris dans le groupe des zygomycètes / sous classe des zygomycétales de la classification traditionnelleRemarque importante
: On a tendance actuellement à les rassembler dans le groupe des
Entomophtoromycotina, d'après les résultats de phylogénie moléculaire.
Grossièrement le groupe des zygomycètes a été restreint et porte
actuellement le nom de Mucoromycotina, avec uniquement des organismes
proches du Genre Mucor ( Mucorales, Endogonales et Mortierellales ). la
sous classe des Zygomycétales ( chitridiomycètes) était différenciée de
celle des Oomycétales par la présence d'un gamète biflagellé au cours du
cycle. Ce critère n'est plus pris en compte car il apparaît primitif et
est successivement apparu et disparu au cours de l'évolution. l'ex
groupe des Oomycétales (Chitridiomycètes ) contient maintenant les :
Chytridiomycota, les néocallimastigomycota, les Blastocladiomycota et
les EntomophtoromycotinaCela représente
plusieurs centaines d'espèces qui sont pour la plupart saprophytes.
Cependant ce groupe contient aussi des parasites de plantes ou de
champignons et des mutualistes de plantes qui forment des mycorhizes.
Les données moléculaires indiquent que ce groupe est monophylétique,
confirmant les données morphologiques et physiologiques. En effet, c'est
à partir de ces champignons que l'on rencontre les thalles végétatifs
typiques des champignons que sont la forme levure unicellulaire et la
forme mycélienne pluricellulaire constituée d'hyphes. Néanmoins,
l'intégration du thalle est limitée:-
les cloisons sont très rares et complètes. Cependant, il existe au
moins chez certaines espèces des mouvements de cytoplasme assurant une
communication primitive. De même, en général, lorsque le mycélium est
blessé le cytoplasme devient très visqueux au point de blessure limitant
les dommages subit par le thalle.- les anastomoses n'existent pas. Il est néanmoins possible d'obtenir des hétérocaryons en mixant ensemble deux mycéliums.De
même, il n'existe pas de vraies fructifications pluricellulaires, sauf
chez quelques endogonales. Leur paroi contient en grande quantité une
forme déacétylée de la chitine: le chitosan, ce qui les distingue des
champignons supérieurs.Des
études sont menées pour déterminer leur potentiel des dans les luttes
contre les insectes mais les résultats sont peu probants pour l'instant
car si ces organismes peuvent contrôler à long terme les populations
d'insectes, ils ne peuvent pas juguler une situation épidémique. Leur
cycle haplobiontique peut ou non présenter une reproduction sexuée et ne
présente pas de zoospore flagellée :Après
la pénétration dans l'insecte via la germination d'une conidie, le
mycélium envahit puis tue son hôte. Souvent, le mycélium est dépourvu de
paroi pour éviter de déclencher les défenses de l'hôte et s'appelle
alors protoplaste (en fait un plasmode). Il se produit ensuite soit un
cycle asexué avec production de spores de type conidie (avec
éventuellement une succession de deux spores, la germination de la
première donnant immédiatement la seconde) soit un cycle sexué avec
production de zygospore. La méiose a probablement lieu dans la
zygospore. Celle-ci perdure dans l'environnement puis germe pour donner
immédiatement une conidie qui est la propagule infectieuse. Outre Outre
les entomophthorales, il existe aussi dans ce groupe des saprophytes et
des parasites d'algues et de vertébrés comme par exemple Olpidium et Basidiobolus.
Les premiers possèdent toujours un stage flagellés alors que chez les
seconds la forme flagellée a aussi disparu (mais cet organisme possède
encore un corps basal). | |
| | | | biologista
Sexe :
Messages : 180
Points : 317
Date de naissance : 07/05/1992
Age : 32
Emploi : Chez mon papa
| | Sujet: Re: Classification.générale (cours) Mer 5 Déc 2012 - 23:34 | |
| 2.5 CLASSE DES ASCOMYCETES2.5.1 Caractères généraux-
Hyphes cloisonnés, structure cellulaire, ce qui implique un seul noyau
par cytoplasme. Il y a cependant lieu de nuancer : voir le cycle de la
Pezize.- Reproduction par asques qui sont des fructifications : une asque est normalement composée de 8 ascospores n ( qui sont des endospores )* Les asques sont fréquemment rassemblées dans des carpophores,
ou elles forment l'hyménium, a noter que le carpophore est le "
champignon" du langage courant et ... culinaire pour autant qu'il
s'agisse de champignos comestibles ; * Endospores = spore se formant dans le cytoplasme d'une cellule.cycle de la PezizeIl s'agit d'un champignon saprophyte forestier.Des
filaments pluricellulaires haploïdes naissent à partir de spores (n).
Prenons le cas d'une espèce hétérothallique : il existe des filament +
provenant de spores + et des filaments - provenant de spores -. NB. Il existe aussi des espèces homothalliques A
la base de ce qui deviendra le carpophore, un gamétange mâle ( ou
anthéridie ) et un gamétange femelle ( ou oogone ) sont formés; le
contenu de l'anthéridie va fusionner avec celui de l'oogone ( en passant
par une sorte de canam de fécondation ou trichogyne ) . A noter que les
gamétanges mâle et femelle sont pluri-nucléés et contiennent des noyaux
n  Il Il
y a aussi lieu de remarquer que les noyaux provenant des deux
gamétanges restent indépendant ,haploïdes , et qu'ils subissent des
caryocinèses conjuguées : ils réalisent ainsi une série de nouveaux
filaments appelés dicaryons . A
ce moment chaque élément du filament est en fait un article binucléé à
noyaux n, l'un venant de la spore +, l'autre de la spore - . Ce mycélium
à dicaryons tapisse l'intérieur du carpophore tandis que l'extérieur de
ce carpophore est composé de filaments uninucléés issus des spores + ou
- A
l'intérieur du carpophore, des cellules en forme de crochet
apparaissent : ce sont les asques dont les deux noyaux diaryons
fusionnent pour donner un zygote (2n), le zygote subit la méiose
immédiatement après sa formation et chaque cellule se divise à son tour
une fois par une cinèse normale ( mitose ). celà donne 8 noyaux (n).
Chaque noyau s'entoure de cytoplasme et d'une membrane et donne une
asquospore n. L'ensemble est logé dans l'asque. Les ascospores seront à
leur tour dispersées. Le carpophore comprend également des filaments
stériles ou paraphyses qui proviennent des filaments de départ ( n ).   Paraphyses + asques = hyménium L'hyménium Paraphyses + asques = hyménium L'hyménium
repose sur le stroma qui présente içi une forme de coupe . La coupe est
l'ensemble de la fructification ou carpophore, ou carpogone.En résumé,
cycle biologique complexe avec une phase n à partir de spores + et de
spores - . Suit une phase (n + n) à dicaryons que l'on peut assimiler
au sporophyte et enfin un retour à la phase n avec spores + et spores -Ce
cycle biologique est parfois ; abrégé; on le rencontre également chez
les Basidiomycètes , mais içi l'asque est remplacé par une baside et
d'autre part la phase à dicaryons est beaucoup plus importante ( le
mycélium et l'entièreté du carpophore sont à dicaryons : à voir au
chapitre suivant )2.5.2 CLASSIFICATION DES ASCOMYCETES2.5.2.1 Classification moderne-Nous voilà devant le groupe de loin le plus important puisqu’il réunit environ les ¾ des espèces connues de champignons.-Il
est caractérisé par l’asque, cellule dans laquelle se déroule la méiose
et la différenciation des ascospores. Typiquement, l’asque contient 8
ascospores (il y a une division supplémentaire après la méiose) mais il
existe des variations par avortement ou divisions supplémentaires.Cette classification est en pleine évolution suite à des travaux basés sur les aspects ultrastructuraux et moléculaires .D’autre
part les différents auteurs ne s’accordent pas toujours sur les niveaux
hiérarchiques et les dénominations des grandes divisions, Classes,
Ordres,…, qui peuvent varier de façon importante. Aussi le tableau
présenté, assez simplifié, est loin d’être ni parfait ni définitif.voiçi la tendance moderne :- Dans
un premier temps on distingue trois grands phylums nettement définis,
les Taphrinomycotina, les Saccharomycotina et les Pezizomycotina. Les Les
Ascomycètes ancestraux auraient été filamenteux et le développement
mycélien aurait été perdu à deux reprises d’une part chez les
Taphrinomycotina (Schizosaccharomycètes) et d’autre par chez les
Saccharomycotina.Taphrinomycotina-Ou Archiascomycètes, Archéoascomycètes. Chez
les Taphrinomycotina, le groupe le plus basal, existent deux types
morphologiques : des asques nus et une croissance unicellulaire
(levures) ou un ascocarpe rudimentaire et une croissance mycélienne.NEOLECTOMYCETES Avec le seul genre Neolecta (3 espèces) ressemblant à un Mitrula.
A été qualifié de « dinosaure » des Ascomycètes (à la taille près et
certainement beaucoup plus ancien !), car selon certains il
représenterait un type basal pour tous les Ascomycètes.TAPHRINOMYCETES Taphrinales Le
cycle de développement comprend 2 phases successives, l’une levuriforme
se développe à la surface des organes végétatifs et se conserve dans
les bourgeons puis au printemps la phase mycélienne, filamenteuse
devient parasite et provoque des déformations importantes sur les
feuilles, les rameaux ou les fruits. Les asques se forment à la
surface des organes attaqués. Taphrina deformans, la cloque du Pêcher, T. pruni, la pochette du prunierT. populina sur Peuplier. Protomyces parfois classé dans les Protomycetales, produit des galles sur de nombreuses herbacées.SCHIZOSACCHAROMYCETES Ce
sont des formes levures qui se distinguent des Saccharomycètes par leur
mode de multiplication par partition cellulaire et non par
bourgeonnement. On connaît 4 espèces de SchizosaccharomycesPNEUMOCYSTIDOMYCETESLes
Pneumocystis sont des parasites unicellulaires des mammifères et, chez
l’homme, à l’origine de graves pneumonies, en particulier chez les
immuodéficients. D’abord considéré comme des protistes, ce sont des
études ultrastructurales puis de biologie moléculaire qui ont permis de
préciser leur place.Saccharomycotina Ou
Saccharomycètes, Hemiascomycètes (p.p.). -Champignons essentiellement
levuriformes rarement filamenteux, à multiplication végétative par
bourgeonnement. Des bourgeonnements successifs peuvent, dans certains,
cas aboutir à des pseudomycéliums. Les asques, isolés, résultent de la
fusion de deux cellules. Un seul ordre : Saccharomycétales.
Beaucoup se développent à la surface des végétaux, en particulier des
fruits, utilisant les exsudats sucrés, telle l’espèce bien connue : Saccharomyces cerevisiae, la levure de Bière domestiquée depuis des siècles.Certains vivent en relation avec les animaux, et nous sommes directement concernés par Candida albicans, hôte fréquent de nos muqueuses qui peut devenir pathogène (muguet). A signaler aussi les Eremothecium, filamenteux, parasites des végétaux dont une espèce, E. gossypii, dangereuse pour le cotonPezizomycotinaOn
les désigne aussi sous le terme d’Euascomycètes. Ce sont les
Ascomycètes filamenteux, ils possèdent souvent des formes de
multiplication asexuée (anamorphes) très diverses et beaucoup d’entre
eux ne sont même connus que par ces seules formes (Champignons
Imparfaits ou Deutéromycètes). Le phénomène de lichénisation, qui
affecte des ordres entiers, a joué un rôle très important dans
l’évolution de ces champigons et s’est produit de façon indépendante
dans plusieurs lignées. Leurs asques sont protégés à l’intérieur
d’organes spéciaux, ascocarpes, de structure variée : périthèce,
apothècie, cleistothèque, ascostroma.PEZIZOMYCETESL’ascocarpe
est une apothècie (ils correspondent à une bonne partie des ex
Discomycètes). Ils comprennent toutes les espèces dont les asques sont
operculés. Mais en dehors de ces formes discoïdes n’oublions pas les
formes hypogées résultant d’une involution de l’apothècie. Cette
transformation s’est produite de façon indépendante dans plusieurs
lignées différentes  coupe dans une apothécieUn seul ordre : Pezizales avec une douzaine de familles. Pezizacées : Peziza, Sarcosphaera Terfezia, Morchellacées : Morchella, Verpa, Disciotis, Discinacées : Gyromitra, Tubéracées : Tuber, Choiroyces, Terféziacées : Terfezia, Tirmania, Helvellacées : Helvella, Sarcoscyphacées : Sarcoscypha, Pyronematacées : Aleuria, Otidea, Caloscyphacées : Caloscypha.ORBILIOMYCETESCe coupe dans une apothécieUn seul ordre : Pezizales avec une douzaine de familles. Pezizacées : Peziza, Sarcosphaera Terfezia, Morchellacées : Morchella, Verpa, Disciotis, Discinacées : Gyromitra, Tubéracées : Tuber, Choiroyces, Terféziacées : Terfezia, Tirmania, Helvellacées : Helvella, Sarcoscyphacées : Sarcoscypha, Pyronematacées : Aleuria, Otidea, Caloscyphacées : Caloscypha.ORBILIOMYCETESCe
sont également des discomycètes, leurs apothècies gélatineuses sont de
petite taille et les asques sont inoperculés. Certains ont des
anamorphes (Arthrobotrys) nématophages.DOTHIDEOMYCETESCorrespondent
en grande partie aux Loculoascomycètes dont l’ascocarpe d’abord
confondu avec le vrai périthèce des « Pyrénomycètes » (rangés dans les
Sordariomycètes) est en fait un stroma dans lequel se creusent
secondairement une ou plusieurs cavités dans lesquelles se développent
les asques. C’est l’ascostroma corrélé à des asques bituniqués, c’est à
dire à paroi constituée de 2 couches séparables.La
distinction entre les divers ordres n’est pas toujours facile, elle est
basée en particulier sur le processus de formation des locules à
l’intérieur de l’ascostroma. Citons :Dothideales : avec le genre Mycosphaerella, plus d’un millier d’espèces, dont beaucoup sont parasites des végétaux. C’est aussi à cet ordre que se rattache Aureobasidium pullulans, forme conidienne très fréquente à l’automne sur les débris végétaux humides.Pleosporales : avec Pleospora, aux spores mûriformes. Venturia agents des tavelures des fruitiers. Herpotrichia qui enveloppe de ses filaments sombres les pousses de conifères enfouis sous les neiges tardives.Capnodiales
: elles sont responsables des « fumagines » recouvrant les organes
chlorophylliens des végétaux d’une couche de mycélium très sombre.ARTHRONIOMYCETES Petit groupe voisin du précèdent. Forment des microlichens surtout dans les régions tropicales.EUROTIOMYCETESRegroupent
des champignons avec des types d’ascocarpes bien différents. D’une part
ceux correspondent aux anciens Plectomycètes. Leurs asques ont une
paroi fine, évanescente et sont dispersés sans ordre apparent dans
l’ascocarpe du type cleistothèque ou à paroi très diffuse. D’autre part
des ascocarpes de type périthèce, s’ouvrant par une ostiole.Eurotiales : Bien connus par leurs formes conidiennes : Penicillium, Aspergillus,...D’où
leur importance économique. Pensez à la Pénicilline ou au Roquefort !
Les stades parfaits sont souvent constitués de cleistothèques à paroi
peu consistante avec des ascospores lenticulaires à épaississement
équatorial. Mais il y a une grande variété de formes et c’est ici que
viennent se placer les « truffes de cerf » Elaphomyces.Onygenales : Nombreuses espèces kératinophiles, donc se développant sur des restes animaux : poils, corne... (Onygena). Mais aussi parasites : diverses mycoses dont les teignes (Trichophyton) et aussi de bien plus dangereuses : histoplasmoses et coccidiomycoses.Pyrenulales, Verrucariales, Chaetothyriales : Champignons lichénisants, classés jadis dans les Pyrénolichens. Verrucaria maura recouvre d’une mince pellicule noire les rochers littoraux dans la zone supérieure de balancement des marées.LEOTIOMYCETESCe
sont principalement des discomycètes avec asques inoperculés à paroi
simple (unituniqués). Certains produisent des ascocarpes charnus tandis
que des espèces parasites ont des fructifications plus discrètes
certaines entièrement closes (Erysiphales). La classification des
Léotiomycètes est encore loin d’être bien établie.Helotiales : On y rencontre des formes charnues comme avec les genres Leotia, Bulgaria, Mitrula, Spathularia et aussi Chlorociboria. Mais les Géoglossacées sont à exclure. Les Sclérotiniacées sont des parasites importants pour l’agriculture, exemples Botryotina avec sa forme conidienne Botrytis qui provoque les pourriture grise, ou Monilina dont la forme conidienne Monilia provoque les pourritures brunes des fruits.Cyttariales Un seul genre, Cyttaria qui possède de curieuses fructifications sphériques, il est parasite des hêtres austraux (Nothofagus) et se rencontre donc uniquement sur le pourtour du pacifique sud, Andes de Patagonie, Nouvelle Zélande, Ouest de l’Australie.. Rhytismatales : les ascocarpes sont inclus dans des stromas immergés dans les tissus de l’hôte. Genre typique Rhytisma (R. acerinum, R. salicinum) taches noires de l’Erable ou du Saule. Erysiphales
: Les ascocarpes entièrement clos ne correspondent ni à la définition
exacte d’un périthèce ni à celle d’une cleistothèque. Le terme de
chasmothèque a été proposé. Ce sont des parasites foliaires, les
Oïdiums, qui s’attaquent à un très grand nombre de plantes. Certains
sont redoutables pour les cultures (vigne, pommier, rosier). Il
représentent peut être une entité bien particulière au sein des
Léotiomycètes.LECANOROMYCETESCette
classe est extrêmement importante par le nombre de ces espèces (plus de
13000) et du fait que la plus grande part sont des champignons
lichénisants ou lichénicoles. Les ascocarpes sont des apothècies
(Discolichens), ils ne sont périthéciens que dans de rares cas. Les
asques inoperculés ont des parois multicouches, dont plus épaisses très
visibles.On distingue plusieurs ordres : Lécanorales, Peltigérales, Ostropales, Umbilicariales…LICHINOMYCETESDiscolichens
à asques inoperculés, un seul ordre : Lichinales. Lichina pygmaea forme
des coussinets denses sur les rochers littoraux.SORDARIOMYCETESEnglobent
l’essentiel des « Pyrénomycètes » dont l’ascocarpe est un périthèce, nu
ou inclus dans un stroma et les asques unituniqués. C’est un ensemble
très important par le nombre de ses espèces, seuls quelques ordres parmi
les plus importants sont cités. Méliolales
: Ce sont des agents de Fumagines. Ectoparasites des organes verts sur
lesquels se développe un mycélium de couleur foncée. Les périthèces se
forment à la surface d’un mince stroma. Ce sont surtout des champignons
de régions tropicales humides. Hypocréales
(comprennent les Clavicipitales) : Les ascocarpes (périthèces) sont
immergés dans un stroma charnu, souvent vivement coloré, ou disposés à
sa surface. Beaucoup sont des parasites de plantes Nectria, Claviceps purpurea (l’ergot du seigle), Epichloë typhina (la quenouille des graminées), d’insectes ou d’arachnides (Cordyceps) ou d’autres champignons (Hypomyces, Cordyceps). Et aussi des formes saprotrophes (Hypocreopsis) Ophiostomatales : Périthèces isolés, asques à paroi évanescente. Ophiostoma ulmi est le responsable de la « maladie hollandaise » de l’Orme. Diaporthales : Les périthèces sont noirs isolés ou groupés et immergés dans un stroma. A noter Cryphonectria parasitica qui provoque le chancre du châtaignier. Sordariales : Nombreuses espèces coprophiles. Neurospora crassa est la « Drosophile » des mycologues généticiens. Xylariales : Périthèces inclus dans des stromas durs, de couleur sombre. Beaucoup sont xylophages : Xylaria, Hypoxylon, Daldinia, Diatripes. …LABOULBENIOMYCETESChampignons Ophiostomatales : Périthèces isolés, asques à paroi évanescente. Ophiostoma ulmi est le responsable de la « maladie hollandaise » de l’Orme. Diaporthales : Les périthèces sont noirs isolés ou groupés et immergés dans un stroma. A noter Cryphonectria parasitica qui provoque le chancre du châtaignier. Sordariales : Nombreuses espèces coprophiles. Neurospora crassa est la « Drosophile » des mycologues généticiens. Xylariales : Périthèces inclus dans des stromas durs, de couleur sombre. Beaucoup sont xylophages : Xylaria, Hypoxylon, Daldinia, Diatripes. …LABOULBENIOMYCETESChampignons
très curieux par leur mode de vie en ectoparasites d’arthropodes. Leur
place dans la classification reste encore à déterminer exactement. Une
seule chose est certaine, ce sont bien des Ascomycètes. Chez certains il
zxiste des individu mâles et femelles séparés, cas très rare chez les
Ascomycètes. Deux ordres , Pyxidophorales et Laboulbeniales.Cette
classification moderne que nous venons d'ébaucher tant à être la
classification officielle et aussi celle qui est utilisée dans le
domaine de la mycologie théorique ou botanique théorique, elle n'est
cependant pas définitive ;La
classification traditionnelle est encore utilisée en agronomie, par des
praticiens et a l'avantage de faire partie d'un langage
traditionnellement utilisé.D'autre
part elle est plus simple et convient pour des identifications plus
générales, de praticiens ou d'amateurs, ne faisant essentiellement appel
qu'à des caractères morphologiques ne nécessitant qu'une loupe ou un
binoculaire. Nous la présentons dans ce qui suit en ne reprenant que les
groupes ayant intérêt à être connus en agronomie ( phytopathologie,
phytopharmacie ...)Cette
classification se ventile en +/- 15.000 espèces groupées en une
quinzaine d'ordre, comme nous l'avons dit nous ne reprenons que ceux qui
intéressent l'agronomie 2.5.2.2 Classification traditionnelle Deux sous classes : - les Hémiascomycètes ou Hémiascomycétales, Ascomycètes à structure très simple, pas de carpophore, asques nues- les Euascomycètes ou Euascomycétales, Ascomycètes typiques avec carpophores volumineuxParmi ces deux sous-classes, l'on distingue plusieurs ordres dont nous donnont les principaux intéressant l'agronomie.Sous-classe des Hémiascomycètes :Ordres des Endomycétales et des Taphrinales ( Exoascales )Sous-classe des Euascomycètes Ordres des Eurotiales ( Plectascales ), des Erysiphales, Hypocréales, Pezizales,TubéralesExemples intéressant l'agronomie :Hémiascomycètes - Ordre des Endomycétales : famille des SaccharomycétacéesAvec essentiellement la genre Saccharomyces ou levures : levure de bière, levure de pain, d'alcool, de vinification.La levure est constituée d'une seule cellule qui se reproduit par bourgeonnement en milieu riche en sucres.  Si le milieu est aérobie, le sucre est oxydé. En boulangerie, la levure fait lever la pâte qui gonfle par libération de CO2 .C6H12O6(sucre) + 6 O2 ---> 6 CO2(gaz dioxyde de carbone) + 6 H2O + 674 CaloriesEn milieu anaérobie, le sucre est transformé en alcool ( = fermentation alcoolique )C6H12O6 ---> 2 CH3CH2OH (alcool éthylique) + 2 CO2 + 26 CaloriesCette dernière réaction est utilisée en distillerie et en brasserie ; Si le milieu est aérobie, le sucre est oxydé. En boulangerie, la levure fait lever la pâte qui gonfle par libération de CO2 .C6H12O6(sucre) + 6 O2 ---> 6 CO2(gaz dioxyde de carbone) + 6 H2O + 674 CaloriesEn milieu anaérobie, le sucre est transformé en alcool ( = fermentation alcoolique )C6H12O6 ---> 2 CH3CH2OH (alcool éthylique) + 2 CO2 + 26 CaloriesCette dernière réaction est utilisée en distillerie et en brasserie ;  Saccharomyces cerevisiae de 6 à 10 microns ( 1 micron = 1/10 millimètre ) Notons Saccharomyces cerevisiae de 6 à 10 microns ( 1 micron = 1/10 millimètre ) Notons
encore que les Levures ont été longtemps classées ailleurs, jusqu'au
moment où l'on s'est aperçu que les cellules de levure peuvent -
exceptionnellement - donner 4 endospores que l'on assimile alors à une
asque. En réalité il s'agit de cellules isolées ou associées en
colonies . Hémiascomycètes - Ordre des Taphrinales- Taphrina . aurea : cause la cloque du peuplier. Boursouflures dorées apparaissant sur les feuilles au début de l'été- Taphrina . pruni : cause de la cloque sur prunier Cloque sur Prunes- exoascus. deformans ( Taphrina deformans ) : cloque du pêcher Cloque sur Prunes- exoascus. deformans ( Taphrina deformans ) : cloque du pêcher   Euascomycètes : Ordre des Eurotiales ( jadis plectascales )Champignons encore primitifs appelés moisissures, produisant des conidies ou exospores. Formes de reproduction sexuée assez rare. Euascomycètes : Ordre des Eurotiales ( jadis plectascales )Champignons encore primitifs appelés moisissures, produisant des conidies ou exospores. Formes de reproduction sexuée assez rare.  Genre Aspergilus : A.niger : aide à la putréfaction; Genre Aspergilus : A.niger : aide à la putréfaction;  Genre Genre
Pénicilium : P. crustaeceum : fromage de Roquefort ; P. notatum, dont
on a isolé la pénicilline : antibiotique actif sur streptocoques et
staphylocoques, voir microbiologie Euascomycètes : Ordre des ErysiphalesParasites
responsables de maladies des végétaux appelées " blanc " ou " oïdium"
ou vrai mildiou ( de meal-dew) sur pommiers, pêcher, rosiers, chênes et
de nombreuses plantes supérieures très éloignées les unes des autres au
point de vue systématique , par exemple les graminée ou Poacées ( Oïdium
sur froment ) .  Cycle général des agents de l'oïdium de la Vigne : Uncinula necator Euascomycètes : Ordre des Hypocréales ( ou Pyrénomycétales )- Cycle général des agents de l'oïdium de la Vigne : Uncinula necator Euascomycètes : Ordre des Hypocréales ( ou Pyrénomycétales )-
Claviceps purpuera : Ergot du seigle : très toxique, responsable de
mortalités importantes dans le passé ( danse de saint-Guy ) - Nectria sp : responsable du chancre du pommier , du peuplier ( chancre sec ), et d'autres essences fruitières ou forestières. Euascomycètes : Ordre des Pezizales ( jadis Discomycètes )Avec la Pezize en exemple, dont le cycle biologique a été décrit plus haut : carpophore en coupe, tapissé intérieurement par l'hyméniumAutres genres - Nectria sp : responsable du chancre du pommier , du peuplier ( chancre sec ), et d'autres essences fruitières ou forestières. Euascomycètes : Ordre des Pezizales ( jadis Discomycètes )Avec la Pezize en exemple, dont le cycle biologique a été décrit plus haut : carpophore en coupe, tapissé intérieurement par l'hyméniumAutres genres
: le pyronema ; les Morilles ; les helvelles ; les Botytis ( B. cinerea
: pourriture grise des fruits, de la vigne, des bulbes ) ; le Monilia (
moniliose des fruits) .Euascomycètes : Ordre des Phacidiales G.Rhytisma, cause les tâches noires des feuilles d'Erables. D'autres Ascomycètes s'attaquent aux tissus animaux et humains :La pelade : chute des cheveuxLe "muguet" : Candida albicans, dans la bouche des enfants, des vieillards.La mycose des nageurs | |
| | | | biologista
Sexe :
Messages : 180
Points : 317
Date de naissance : 07/05/1992
Age : 32
Emploi : Chez mon papa
| | Sujet: Re: Classification.générale (cours) Mer 5 Déc 2012 - 23:43 | |
| 2.6 CLASSE DES BASIDIOMYCETES2.6.1 Caractères générauxCe sont
généralement les "champignons" du langage courant : hyphes cloisonnés de
structure articulaire ; fructifications ou carpophores contenant des
basides. ( chez les Ascomycètes, les carpophores contenaient des asques
)  La baside et les basidiospores se présentent généralement La baside et les basidiospores se présentent généralement
comme suit : Baside = élément binucléé ( deux noyaux n ) dont les
élément fusionnent à un moment donné ( noyau 2n = zygote ) à partir du
dicaryon de la baside. Viennent ensuite les cinèses réductionnelles,
donnant 4 noyaux n . Chaque noyau s'isole extérieurement à la baside, au
sommet d'un pédicelle et devient une basidiospore qui est donc une
exospore. Comme chez les
Ascomycètes, le mycélium est constitué au départ par deux filaments n (
il y a généralement hétérothallie ) . Ces filaments vont fusionner et
deviennent des hyphes à dicaryons des hyphes à dicaryons. La phase à
dicaryons est beaucoup plus longue chez les Basidios que chez les Ascos :
elle constitue l'essentiel du mycelium et du carpophore ( pied, chapeau
avec son hymenium, anneau, volve, voile ).  Chez les Chez les
basidiomycètes, comme les coprins par exemple, la génération
gamétophytique est représentée par des filaments mycéliens haploïdes.
Ces filaments (= le mycélium primaire) réalisent la reproduction sexuée
en s'unissant deux à deux, c'est une somatogamie ou périttogamie.Dans un premier temps, seuls les cytoplasmes des cellules fusionnent, on dit qu'il y a plasmogamie. Ceci donne naissance à un mycélium dicaryotique
(=mycélium secondaire), c'est à dire constitué de cellules à deux
noyaux haploïdes. Cette phase dicaryotique spécifique des champignons
constitue la génération sporophytique. Le mycélium dicaryotique
s'organise en pseudotissus et forme le carpophore.Au niveau des sporocystes, a lieu la fusion de deux noyaux haploïdes, ou caryogamie,
bientôt suivie d'une méiose qui génèrera quatre spores méiotiques
exogènes. Les sporocyte sont des basides et les spores méiotiques des
basidiospores. Ces spores seront à l'origine du mycélium gamétophytique
haploïde et le cycle est bouclé.  Notons encore Notons encore
que si le reproduction sexuée ( par le carpophore ) permet d'établir le
cycle complet des champignons , il existe également une multiplication
végétative par bourgeonnement ; cette dernière serait la plus fréquente.C’est
l’ensemble dans lequel nous allons trouver la grande majorité des
champignons qui intéressent les mycophages et les mycologues amateurs et
qui sont les mieux connus. Cependant pour ces derniers qui ont déjà
acquis de bonnes connaissances de leur systématique il va falloir
réviser quelques certitudes !En premier lieu
parce que il y a de multiples convergences de formes, de sorte que les
termes classiques, Hyménomycètes, Aphyllophorales, Gastéromycètes tout
comme « Clavaire », « Hydne », « Polypore », ..., n’ont plus qu’une
valeur physionomique. De même que les distinctions entre
Phragmobasidiés, Hétérobasidiés, Homobasidiés sont moins strictes que ce
que l’on pensait . Comme si, dès le début de leur évolution, les
Basidiomycètes ancestraux possédaient en puissance toutes ces
possibilités morphologiques qui ont pu se révéler à plusieurs reprises
et de façon indépendante au cours de leur différenciation.Et tout d’abord
où se situe l’origine des Basidiomycètes ? La question ne paraît pas
résolue. La plupart des analyses phylogéniques montrent un enracinement
commun avec les Ascomycètes quelque part dans les Zygomycètes. La seule
chose qui paraisse certaine est leur monophylie, c’est à dire une
origine commune aux deux embranchements actuels. Ce qui sépare en
premier lieu les deux groupes est le type de cellule qui produit les
spores reproductives, ici c’est donc la baside qui produit des
basidiospores externes, 4 presque toujours. A la différence de l’asque
qui ne montre que des variations de forme réduite, la baside présente
des types morphologiques très variables. Différents types de basides. 1 : Holobaside d’Agaricale. 2 : Holobaside réduite de Calocera. 3 : Hétérobaside de Tulasnella avec épibasides. 4 : Hétérobaside de Tremella incomplétement divisée. 5 : Phragmobaside d’Auricularia. 6 : Phragmobasides de Puccinia issues d’une téliospore (probaside)2.6.2 CLASSIFICATION GENERALE DES BASIDIOMYCETES2.6.2.1 Classification moderneTout comme pour les ascomycètes, une classification moderne, basée sur la phylogénie s'ébauche, en voiçi la structure : Différents types de basides. 1 : Holobaside d’Agaricale. 2 : Holobaside réduite de Calocera. 3 : Hétérobaside de Tulasnella avec épibasides. 4 : Hétérobaside de Tremella incomplétement divisée. 5 : Phragmobaside d’Auricularia. 6 : Phragmobasides de Puccinia issues d’une téliospore (probaside)2.6.2 CLASSIFICATION GENERALE DES BASIDIOMYCETES2.6.2.1 Classification moderneTout comme pour les ascomycètes, une classification moderne, basée sur la phylogénie s'ébauche, en voiçi la structure :  Voir le site de l'association mycologique de Toulouse : http://www.associationmycologiquetoulouse.ups-tlse.fr/spip.php?article33Pucciniomycotina(Uredinomycotina) Plus Voir le site de l'association mycologique de Toulouse : http://www.associationmycologiquetoulouse.ups-tlse.fr/spip.php?article33Pucciniomycotina(Uredinomycotina) Plus
de 8000 espèces décrites, dont la majeure partie appartiennent à
l’ordre des Pucciniales (les Rouilles des végétaux). En dehors des
phytoparasites, il existe aussi des mycoparasites, des parasites
d’insectes et des saprotrophes. Certaines espèces sont levuriformes, la
présence de basidiocarpes est exceptionnelle.PUCCINIOMYCETES (Urédinomycètes)Ce sont tous des parasites, ils possèdent une phragmobaside le plus souvent issue d’une téliospore.Pucciniales (Urédinales).Les
Rouilles des végétaux ont un cycle complexe dans lequel, en plus de la
forme parfaite (téliospores), on peut observer des organes sexués
(spermogonies) et des anamorphes de multiplication, successivement :
écies (écidies) donnant les éciospores puis urédo produisant les
urédospores. La baside naît d’une téliospore qui joue souvent un rôle de
conservation (voir figure basides n°6). Beaucoup sont hétéroxènes,
c’est à dire qu’elles accomplissent leur cycle sur 2 plantes appartenant
à des familles différentes. L’exemple classique est celui de la rouille
noire du blé, Puccinia graminis dont la phase écidienne est hébergée par des Berberis et la phase urédotélienne par des Graminées.Ce
cycle peut être plus ou moins réduit par disparition d’une ou plusieurs
phases jusqu’à ne comporter que des téliospores comme chez Puccinia malvacearum. Les rouilles ont une grande importance économique en raison des dégâts qu’elles provoquent sur les cultures, céréales (Puccinia), légumineuses (Uromyces), florales (Phragmidium sur rosiers), caféier (Hemileia)...voir cycle dans classification traditionnelleTrois autres ordres sont à citer pour leur curieuse biologie.Helicobasidiales Ont d’abord une phase haploïde parasites dans les écies de Pucciniales (Tuberculina) et après fécondation. sur les racines de plantes pendant leur dicaryophase.Septobasidiales
Elles vivent en symbiose avec des cochenilles. Elles forment des
croûtes à la surface des écorces, abritant ainsi des colonies de
cochenilles dont elles parasitent un certain nombre d’individus. Platygloelales A noter encore avec Eocronartium et Jola parasites des mousses aux minuscules fructifications clavarioïdes. CYSTOBASIDIOMYCETES, AGARICOSTILBOMYCETESCes classes réunissent essentiellement des champignons totalement ou partiellement levuriformes.MICROBOTRYOMYCETESPrincipalement représentés par les Microbotryales,
phytoparasites dont beaucoup d’espèces se localisent dans les anthères
des fleurs. Etaient classées avec les Ustilaginales (les Charbons) en
raison de leurs téliospores sphériques, plus ou moins ornementées
ressemblant à celles de ces champignons. Leurs basidiospores germent en
donnant une forme levure. Exemple Microbotryum violaceum = Ustilago violaca dans les anthères de certaines Caryophyllacées.MIXIOMYCETES Une seule espèce, Mixia osmundae, qui mérite d’être citée car elle a d’abord été décrite comme un Ascomycète. Ustilaginomycotina Réunissent
près de 1500 espèces de parasites des végétaux. Beaucoup possèdent
des téliospores et une phase haploïde levuriforme.USTILAGINOMYCETESCe sont les « Charbons » dont l’importance économique est loin d’être négligeable, charbon du blé (Ustilago tritici), du maïs (Ustilago maydis
) par exemple. Leur sporulation constitue des amas (sores) de
téliospores de couleur foncée. Ces spores sont libérées par éclatement
des tissus de l’hôte. Leur germination donne une phragmobaside. Deux
ordres :Ustilaginales surtout parasites sur Graminées et Cypéracées.Urocystales sur diverses Mono et Dicotylédones (Urocystis ficariae sur Ranunculus ficaria).EXOBASIDIOMYCETESSe différencie des précédents par la prèsence d’une holobaside. On y trouve des champignons produisant ou non des téliospores.Tilletiales Avec des téliospores de couleur sombre qui, pour la plupart, se développent dans les ovaires de Graminées (Tilletia caries, la carie du blé).Entylomatales,
les téliospores sont peu ou pas colorées et dispersées dans les
parenchymes foliaire de l’hôte. Exemple le très commun Entyloma ficariae.Doassansiales
Elles se distinguent principalement des précédentes par l’agglomération
des téliospores en masse. Elles parasitent surtout des plantes
aquatiques.Exobasidiales
Il n’y a pas de téliospores et les basides sont produites le plus
souvent à la surface des organes parasités, feuilles principalement, qui
sont plus ou moins profondément déformés, l’exemple type est Exobasidium rhododendri qui produit de grosses galles vivement colorées sur le feuillage des Rhododendrons.Microstromatales et Georgesfischeriales ne regroupent qu’un petit nombre d’espèces.ENTORRHIZOMYCETESGroupe très réduit (un seul genre, Entorrhiza)
qui se distingue par des téliospores se différenciant à l’intérieur
des cellules vivantes dans les tissus des racines de l’hôte . Ces
téliospores germent sans donner de baside extérieure mais en se
cloisonnant et seuls 4 stérigmates émergent. Leur rattachement
traditionnel aux Ustilaginomycotina, basé sur leur parasitisme, est
douteux, certaines analyses les font apparaître à la base de l’ensemble
des Basidiomycètes.AgaricomycotinaIls
comprennent le tiers des espèces décrites de champignons et l’énorme
majorité de celles qui intéressent le plus les mycologues amateurs. A
part quelques unités, c’est parmi eux que l’on trouve les espèces
comestibles, et aussi des plus toxiques. De plus leur importance
écologique et économique est considérable. En tant que mycorhiziens ils
contribuent à la croissance et au maintien d’une grande partie des
forêts mondiales et par là à la production de biomasse, bois en
particulier. Bois que bon nombre d’autres se chargent ensuite de
détruire. Quelques font l’objet d’une culture et d’un commerce
intensifs. Leur classification traditionnelle basée sur la structure de
la baside (hétéro ou holobaside), le type de basidiocarpe, d’hyménium...
a été profondément bouleversée. Les recherches phylogénétiques basées
aujourd’hui non plus sur un seul mais sur plusieurs fragment de génome
donnent des résultats relativement solides. Il semble donc que l’on
puisse accepter trois ensembles (classes) d’importances inégales,
Agaricomycètes, Dacrymycètes et Trémellomycètes.TREMELLOMYCETESC’est
la classe qui paraît la plus proche de la souche commune des
Basidiomycota. Les basides sont tétracellulaires avec des cloisons plus
ou moins complètes disposées longitudinalement ou obliquement (voir
figure basides n°4). Les basidiocarpes (quand ils sont présents) sont
gélatineux. La phase haploïde est levuriforme.Trémellales sont typiquement représentées par l’espèce commune Tremella mesenterica, saprotrophe sur bois mort. Beaucoup d’espèces bien moins spectaculaires sont des mycoparasites d’autres Basidiomycètes.Filobasidiales ne produisent pas de basidiocarpe. Ces champignons sont surtout connus par leurs formes levures : Cryptococcus. L’une d’elle, C. neoformans, est un pathogène humain, très dangereux pour les immunodéficients.DACRYMYCETESPetit
ensemble de d’espèces à basidiocarpes gélatineux de faible taille. La
baside est unicellulaire prolongée de deux longs srérigmates qui lui
donnent un aspect fourchu (voir figure basides n°2), holobaside,
hétérobaside ? C’est comme l’on voudra ! Ce sont des décomposeurs de
cellulose (pourritures brunes).Un seul ordre, Dacrymycétales, exemples : Dacrymyces, Calocera.AGARICOMYCETESParmi
tout une série d’ordres dont les relations ne sont pas encore bien
clairement définies se détachent deux ensembles auxquels on donne le
rang de sous-classes, les Agaricomycétidées et les Phallomycétidées.
Restent un certain nombre d’ordres pour lesquels il est encore difficile
d’assigner une place exacte dans une phylogénie générale. Un ensemble
se trouve près de la racine des Agaricomycètes. Ce sont d’une part des
hétérobasidiés, Sébacinales et Auriculariales et d’autre part des
holobasidiées, Cantharellales.Sébacinales
aux basidiomes réduits ou nuls avec basides renflées, subdivisées
longitudinalement et surmontées de 4 épibasides allongées.. Elles jouent
un rôle, longtemps méconnu mais important, dans la mycorhization de
nombreuses plantes comme les Orchidées ou les Ericacées.Auriculariales, elles possèdent des basidiocarpes gélatineux, leurs basides sont typiquement cloisonnées transversalement Les genres Auricularia et Exidia sont bien connus.Cantharellales, homobasidiées, comprennent les Cantharellus, Craterellus et aussi Hydnum, Clavulina. Le sort des Tulasnellales,
aux basides de forme particulière (figure basides n°3), n’est pas
résolu, pour les uns cela reste un ordre distinct alors que certaines
analyses les incluent dans les Cantharellales.Le
deuxième ensemble est constitué de champignons homobasidiés, très
souvent lignicoles aux basidiomes de consistance coriace, parfois
réduits « croutes ». On peut citer :CorticialesPolyporales : Polyporus, Fomitopsis, LaetiporusHyménochaetales : Coltricia, Inonotus, PhellinusTéléphorales : TelephoraPHALLOMYCETIDEESElles
ne correspondent pas exactement aux ex Gastéromycètes car ce
regroupement concerne aussi des champignons qui n’y étaient pas inclus
et en exclu au contraire d’autres.Phallales. Les basidiospores baignent dans une gléba mucilagineuse à odeur puissante (dispersion entomophile) (Phallus, Mutinus, Clathrus...)Gomphales. Réunissent avec les Gomphus, des types aux morphologies très diverses : coraloïde (Ramaria), en massue (Clavariadelphus), hypogée (Gautiera)...De plus deux nouveaux ordres sont crées :Géastrales pour les familles des Géastracées, Sclerogastracées et SphaerobolaceaeHystérangiales, essentiellement hypogées.AGARICOMYCETIDEESNous voici au cœur de l’essentiel de la matière des atlas de terrain, de ce qui intéresse le plus l’amateur de champignons.Russulales.
Suivant les auteurs cet ordre est souvent considéré comme un groupe
frère des autres Agaricomycetidées. La famille des Russulacées est
évidemment bien connue : Russula, Lactarius
constituent une part importante de la flore de nos sous bois. Oui mais,
certaines espèces tropicales sont annelées ou ressemblent à des
pleurotes tandis que des formes souterraines (Gymnomyces, Macowanites, Zelleromyces) viennent se loger au sein même de ces deux genres qui sont eux mêmes paraphylétiques. En effet les Russules du sous-genre Compacta sont plus proches des Lactaires blancs (L. piperatus
...) que des autres Russules. Et puis la biologie moléculaire a révélé
des parentés à priori peu évidentes avec d’autres familles :
Peniophoracées, Auriscalpiacées, Hericiacées, Stéréacées.... Ainsi sont
inclus dans les Russulales des genres comme Albatrellus, Amylostereum, Artomyces, Auriscalpium, Hericium, Heterobasidion, Peniophora, Stereum.
A noter que ces regroupements ne correspondent pas uniquement à des
données biomoléculaires, on trouve de façon presque générale chez ces
champignons des spores amyloïdes et des lacticifères.Agaricales.
C’est l’essentiel des champignons lamellés mais pas seulement.car il y a
aussi des formes gastroïdes ou clavarioïdes.. On peut y distinguer 5
clades (rameaux) principaux : Plutéoïdes : Plutéacées, Amanitacées, Pleurotacées. Hygrophoroïdes : Hygrophoracées, Ptérulacées, Typhulacées. Marasmioïde : Marasmiacées, Omphalotacées, Schizophyllacées... Agaricoïdes : Agaricacées (inclus Lycoperdon),
Strophariacées, Inocybacées Cortinariacées... et d’autres formes
gastroîdes : Nidulariacées, Hymenogastracées. Tricholomatoïdes :
Tricholomatacées, Lyophyllacées, Entolomatacées, Mycenacées...Bolétales.
On sait déjà qu’ils ne s’agit pas uniquement de formes charnues à
hymenium poré. Ici aussi la biologie moléculaire apporte des précisions
mais qui méritent encore d’être approfondies. On distingue un certain
nombre de lignées bien individualisées (sous-ordres ou familles) :
Boletinées, autour des genres Boletus, Xerocomus, Porphyrellus, Chalciporus, Phylloporus... Suillinées : Suillus, Gomphidius Rhizopogon... Paxillinées : Paxillus, Melanogaster... Sclerodermatinées : Gyroporus, Scleroderma, Calostoma, Pisolithus. Hygrophoropsidacées : Hygrophoropsis. Serpulaceae : Serpula Coniophoracées : Coniophora.2.6.2.2 Classification traditionnelleComme argumenté dans le chapitre relatif aux ascomycètes, cette classification se révèle toujours très utile, surtout à des fins pratiques.I. Absence de carpophoreBasides cloisonnées. Champignons parasites----------------> Hémibasidiés (ou Hémibasidiomycètes)II. Carpophores généralement présents a) Carpophores gélatineux et basides cloisonnées, Champignons saprophytes----------------> Protobasidésb) Carpophores non gélatineux et basides unicellulaires----------------> Autobasidiés2.6.2.2.1 LA SOUS CLASSE DES HEMIBASIDIESCes champignons généralement parasites sont considérés comme de redoutales ennemis de nombreuses plantes cultivées Du point de vue agronomique, cette sous-classe comporte deux Ordres importants :- Ordre des Ustilaginales- Ordre des Uredinales A) O. Ustilaginales Parasites des céréales, détruisant entièrement les organes floraux Les
ustilaginomycètes regroupent environ 1300 espèces qui sont des
parasites de plantes à fleurs avec un seul hôte et sont donc comme les
rouilles (pucciniomycètes) des pathogènes d'importance économique.
D'autant plus qu'ils attaquent les fleurs et empêchent donc la formation
des graines. A la fin de l'infection, ils forment souvent des amas de
spores noires d'où le nom de charbon qui leur a été donné. Ils attaquent
leur hôte de manière particulière en secrétant via des vésicules des
dépôts à l'intérieur des cellules. Ils alternent souvent entre une phase
mycélienne infectieuse et une phase levure saprophyte. Leurs pores
septaux sont plus simples que chez les agaricomycotina et ne sont pas
des dolipores caractéristiques. Deux grands groupes sont reconnus sur la
base des données moléculaires.- Genre Ustillago exemple U.Tritici C'est
le "charbon" du Froment. Des spores noires germent avec les grains de
pollen sur stigmates des fleurs : le mycélium envahit les ovules, puis
ultérieurement les graines qui en sont issues sans leur causer de
dommages. Ce
mycélium dormant se réveille avec la germination du grain au printemps
suivant et envahit toute la plante issue du grain, détruisant les épis
qui sont alors remplacés par des amas de spores.L'attaque se produit donc en seconde généralement.Les
agriculteurs produisent de moins en moins leur semences, la maladie se
raréfie donc. Traditionnellement on luttait contre cette maladie en
traitant les semences à l'eau chaude sans en altérer les propriétés
germinatives.  ou encore U. maydisL'espèce la plus connue de ce groupe actuellement est Ustilago maydis ou encore U. maydisL'espèce la plus connue de ce groupe actuellement est Ustilago maydis
qui attaque le maïs provoquant des galles (c'est à dire des tumeurs où
le tissu de l'hôte est hypertrophié et mélangé avec le mycélium) qui
remplace les grains dans l'épi. Le génome de ce champignon est
maintenant complètement séquencé. Son analyse devrait permettre de mieux
comprendre sa biologie. Le cycle est le suivant :  Le cycle commence par la germination de la spore diploïde (téliospore) au printemps qui s'accompagne de la méiose. Ceci donne naissance à un promycélium de 4 cellules haploïdes. Chacune de ces cellules produit au moins un bourgeon (le sporidium) Le cycle commence par la germination de la spore diploïde (téliospore) au printemps qui s'accompagne de la méiose. Ceci donne naissance à un promycélium de 4 cellules haploïdes. Chacune de ces cellules produit au moins un bourgeon (le sporidium)
qui est l'équivalent de la basidiospore. Ce processus peut se répéter
plusieurs fois. Si des cellules compatibles se rencontrent, il y a
fusion cellulaire et formation du dicaryon. Ceci entraîne la formation
du mycélium et l'envahissement de la plante (qui ne se produit donc que
s'il y a fusion sexuelle). Lorsqu'il atteint la surface de la plante, le
mycélium peut aussi produire des spores dicaryotiques qui assurent une
propagation asexuée. La fusion nucléaire intervient dans les téliospores
en formation pendant l'automne dans des galles qui occupent la place
des grains dans l'épi, terminant le cycle et assurant la production
d'une forme de résistance pendant l'hiver; - Genre Tilletia : exemple T. tritici C'est la "carie du froment".  Une Une
basidiospore adhérant au grain infecte la toute jeune plante. A la
floraison, l'ovaire est dévoré par le champignon et est remplacé par un
amas de spores binucléées à membrane cellulosique très résistante et
appelées chlamydospores.
Ces spores tombent au sol. A l'automne, fusion des deux noyaux et
formation du zygote. Au printemps suivant interviennent des cinèses
réductionnelles. Les quatre petites cellules n obtenues forment de
petits diverticules dans lesquelles s'engage chaque noyau = quatre
basidiospores.  B) O. UredinalesParasites responsables des maladies connues sous le nom de " rouilles " : voir ci-après. Attaquent des milliers d'espèces de Phanérogames et surtout des graminées sociales ( exemples : céréales ) et surtout des graminées sociales ( exemple : céréales ).En B) O. UredinalesParasites responsables des maladies connues sous le nom de " rouilles " : voir ci-après. Attaquent des milliers d'espèces de Phanérogames et surtout des graminées sociales ( exemples : céréales ) et surtout des graminées sociales ( exemple : céréales ).En
conditions d'années humides, les dégâts peuvent être énormes, par action
sur les feuilles et développement de plusieurs types de spores dont un
type de teinte jaune orange qui leur vaut leur nom. Le cycle biologique est complexe, passant par un hôte intermédiaire.Exemples :- Puccinia graminis sur froment et épine vinette ( hôte secondaire ) Pustules sur feuilles d'épine vinette- Filament n *Picnides ---------> picnospores , face supérieure--------> cycle végétatif sur épine vinette------>picnides* - Filament n*Ascidies ----------> ecidiospores , face inférieure( 2 noyaux)---------> infestation du plant de froment par les feuilles--------> **Pustules jaune or sur feuilles de froment**------> Pustules sur feuilles d'épine vinette- Filament n *Picnides ---------> picnospores , face supérieure--------> cycle végétatif sur épine vinette------>picnides* - Filament n*Ascidies ----------> ecidiospores , face inférieure( 2 noyaux)---------> infestation du plant de froment par les feuilles--------> **Pustules jaune or sur feuilles de froment**------>
dicaryons dans tissus froment---> pustules jaune or( conidies-
urédospores) - infestation d'autres plants de froment pas autre type de
spore binucléée (conidies --urédospores)---------->
dicaryons------>Téleutospores ( noires de survie )-------->
caryogamie ------->zygote ---->
méiose----->basides-->basidiospores-------------> retour sur
feuilles épine vinette -------- filament n glossairemicromycetes.pdf rouillessynthese.pdf- Melampsora larici-populina sur peuplier et mélèze- Melampsora allii-populina sur peuplier et Allium- Autres Melampsoras avec d'autres hôtes- Gymnosporangium sabine = rouille grillagée du poirier ( Téleutospores sur Genévrier )-Autres glossairemicromycetes.pdf rouillessynthese.pdf- Melampsora larici-populina sur peuplier et mélèze- Melampsora allii-populina sur peuplier et Allium- Autres Melampsoras avec d'autres hôtes- Gymnosporangium sabine = rouille grillagée du poirier ( Téleutospores sur Genévrier )-Autres
rouilles : du Caféier, de la Betterave ( autoïque ), des Graminées de
prairies; des Légumineuses; rouille " jaune " du froment, de l'orge, du
seigle; rouille "couronnée" de l'avoine; des Malvacées, du rosier,
etc...2.6.2.2.2 SOUS-CLASSE DES PROTOBASIDIES Champignons xylophages, exemples : Tremelles ( oreilles de judas ), Hirnéoles  Trémelles sur bois Caractéristique de l'"oreille de Judas"Fructification: Trémelles sur bois Caractéristique de l'"oreille de Judas"Fructification:
sessile ou à pied rudimentaire, 2-8 cm de large et jusqu'à 4 cm de
projection, en forme de bonnet, de coquille ou d'oreille, fixée
latéralement au substrat ou par le sommet; à chair mince,
gélatineuse-caoutchouteuse au frais, devenant cornée et dure en
séchant, pouvant reprendre sa consistance initiale en présence d'eau, à
odeur faible et à saveur agréable.Face supérieure: stérile, habituellement veinée et côtelée, veloutée-tomenteuse, brun pâle, brune ou brun rougeâtre, noircissant en séchant.Face inférieure: fertile, lisse à légèrement plissée, brun jaunâtre, brun rougeâtre, brun grisâtre ou brun pourpré.Spores: allantoïdes, lisses, hyalines, guttulées, 12-16 x 4-7 µm. Sporée blanche.Habitat:
Habituellement en groupes sur les troncs morts de feuillus et de
conifères, en été et en automne. Les individus qui ont poussé tard à
l'automne peuvent se réhydrater après la fonte des neiges printanière et
persister ainsi quelques temps avant de se décomposer. CommunComestibilité: Comestible.2.6.2.2.3 SOUS-CLASSE DES AUTOBASIDIES Ce
sont les " champignons" du langage courant, mais dont le profane ne
retient, en fait, que l'élément extérieur, c'est à dire le carpophore.La
plupart des autobasidiés sont saprophytes, ou parasites des parties
ligneuses des plantes. Beaucoup d'entre eux vivent en symbiose avec les
arbres ( et les végétaux supérieurs ) : leur mycelium peut constituer
des des associations appelées mycorhises avec les racines. Ces
associations sont à bénéfice réciproque ( voir la rubrique pédologie ). une
majorité de nos plantes forestières forment des mycorhises avec
certaines espèces de champignons; Beaucoup de plantes herbacées liées
aux mycorhyses, certaines le sont obligatoirement ( les Orchidées
doivent être associées aux champignons du genre Rhizoctonia ). On admet
de plus en plus que les associations nutritives plantes-microflore
apportent à l'hôte, non seulement des matières alimentaires, mais aussi
des facteurs de croissance et de résistance au parasitisme animal et
végétal. En d'autres termes une mycorhisation abondante serait un moyen
de lutte préventif à l'égard de certaines maladies.Parmi les Autobasidiés, on distingue :- les Hymnomycètes : champignons à hyménium nu, étalé à l'air libre.Exemple : Agaric, champignon blanc de prairie, le plus connu et le plus consommé. - les Gastéromycètes : l'hyménium est enfermé dans le carpophore ( Exemple : la Vesse de loup ou Lycoperdum).A . Les HyménomycètesSouvent saprophytes, vivant sur matière organique abondante ( litière, praiie, bois mort...)Au
printemps une basidiospore germe ---> mycelium qui prolifère tout
l'été; vers l'automne apparaît un carpophore sur le feutrage mycélien
qui provient en réalité de deux basidiospores n de polarité sexuelle
différente ( = hétérothallie )La plus grande partie du mycelium est à dicaryons (plasmogamie avec deux noyaux : n+ et n-
) . La fusion des cytoplasmes et la formation de dicaryons se réalise
très tôt ; le mycélium des Basidiomycètes est constitué dans sa
quasi-entièreté par des dicaryons ( ce qui le distingue des Ascomycètes
ou la dicaryophase se limite à l'intérieur du carpophore ).Un
fait important à signaler dans le développement du mycélium de
Basidiomycètes est la division des articles binucléés par voie d'anses
d'anastomoses.  figure Wikipedia : anses d'anastomoseLe carpophore se compose d'un pied ou stipe, d'un chapeau, d'un anneau, parfois d'un voile. figure Wikipedia : anses d'anastomoseLe carpophore se compose d'un pied ou stipe, d'un chapeau, d'un anneau, parfois d'un voile.  Pas Pas
de croissance polarisée par une initiale. Pourquoi se redressent-il ?
C'est dans la baside que se réalise la fusion nucléaire ( zygote ) puis
la réduction chromosomiqueDIFFERENCES GENERALES ENTRE ASCOMYCETES ET BASIDIOMYCETES| ASCOMYCETES | BASIDIOMYCETES | présence d'organes sexuels ( anthéridies et oogones) à la base du carpophore
| Plus d'organes sexuels : une cellule indifférenciée d'un hyphe n fait office
d'un hyphe + et d'un hyphe -
| | dicaryophase brève : réduite aux hyphes ascogènes ) | la dicaryophase concerne la majeure partie du mycelium et tous les carpophores
| | Deux sexes ( spores + et spores - ) | souvent 4 sexes (= tétrapolarité sexuelle) : les 4 basidiospores d'une même
baside ne sont fécondables qu'avec les éléments d'une autre baside
| "anse de panier" au niveau de la cellule mère au niveau de la cellule-mère de l'asque
| anses d'anastomoses partout dans le mycelium | | la sporée est extérieure au carpophore ( par projection ) | La sporée est intérieure au carpophore | | les filaments mycéliens ne sont jamais binucléés | les filaments mycéliens sont binucléés dans leur immense majorité |
IMPORTANCE DE CE GROUPE DE CHAMPIGNONS1) Nombre d'entre eux sont des champignons dont le carpophore est utilisé commealiment.
Mais il faut distinguer les champignons comestibles (Agaric, Girolle),
les champignons indigestes ( certaines Russules, Bolets, Lactaires ) et
lesn champignons toxiques ( certaines Amanites, dont A. banche, A .
phalloïde, A. vireuse sont mortelles ).La mort est due à une destruction des globules rouges du sang . 2 ) Beaucoup de Basidiomycètes ( et d'Ascomycètes ) sont des saprophytes
, ils débarrassent la surface du sol de nombeux déchets organiques qui
s'y accumulent. A cet égard, les champignons jouent un rôle important
dans les chaînes trophiques. De nombreux noms doivent être cités : Bolets, Fomes,Merulius ( le mérule des bois ).3)
Il existe aussi , parmi les Basidiomycètes, de nombreux parasites (
Rouilles, Charbons, caries, Armillaire, Polyphores, Pleurottes, Coprins
,... )4 ) Il faut encore citer de nombreux basidiomycètes comme mycorhises .5) A la limite, il est évident que certains champignons peuvent être classés comme parasites ou saprophytes. B. Les gastéromycètesHyménium enfermé dans son carpophore.Exemple, Lycoperdum ou saprophytes .LES FUNGI IMPERFECTI Ce sont les champignons dits " imparfaits", encore appelé Adélomycètes, Deutéromycètes ou encore Hyphomycètes .Champignons avec
un mycélium cloisonné, mais dont les fructifications ne sont pas
connues.C'est donc par ignorance qu'ils sont qualifiés d'imparfaits et
beaucoup d'Ascomycètes et de Basidiomycètes étaient jadis rangés dans ce
groupe résiduel.Leur
reproduction se fait par des conidies sur conididiophores. Ceux-çi sont
soit associés dans des conceptacles globuleux appelés pyenides ou étalés
en coussinets appelés acervules ou encore sont indépendants. Trés
nombreux genres ( +/- 1000) et plusieurs milliers d'espèces. Plusieurs classes : Mycelia Sterilia, Coelomycètes, Hyphomycètes, Blastomycètes Le sol
constitue un réservoir inépuisable de champignons imparfaits. Certaines
espèces sont de véritables champignons saprophytes du sol, où ils
accomplissent tout leur cycle de développement ; ils jouent un rôle
important dans la décomposition des débris organiques. D'autres sont des
parasites facultatifs des plantes supérieures, susceptibles de végéter
pour un temps dans le sol ; ils constituent un foyer d'infection pour
les jeunes pousses, et imposent la pratique de l'assolement. D'autres
enfin sont des champignons des litières, localisés dans la couche
humique du sol ; on y rencontre en particulier des espèces prédatrices (Arthrobotrys, Dactylaria),
remarquablement adaptées à la capture des nématodes ou des rotifères.
La mycoflore du sol est le siège de compétitions et d'antagonismes,
manifestés par la sécrétion de toxines ou par le mycoparasitisme, qui
tendent à l'éviction des espèces parasites et contribuent à l'équilibre
de la biocénose.Beaucoup de spores de champignons se retrouvent, à l'état viable, dans l'atmosphère ; elles peuvent être à l'origine de réactions allergiques.Les champignons du sol et de l'air sont les principales sources de contamination par les moisissures
; les dégradations causées par ces micromycètes aux biens de
consommation, produits alimentaires, matériaux et objets fabriqués font
subir de graves préjudices à l'économie.Beaucoup de champignons parasites des plantes cultivées appartiennent aux diverses classes de Fungi imperfecti
: Sphæropsidales (septorioses, rot blanc de la vigne) ; Mélanconiales
(anthracnoses ou pourritures sèches des fruits et légumes, maladie
criblée des arbres fruitiers) ; Hyphomycétales (cercosporioses,
alternarioses, fusarioses, et de nombreux parasites secondaires ou de
faiblesse). Les parasites appartenant à d'autres groupes (Ascomycètes)
ont souvent une phase conidienne très active qui assure la dispersion
locale du champignon pendant la période de végétation. | |
| | | | biologista
Sexe :
Messages : 180
Points : 317
Date de naissance : 07/05/1992
Age : 32
Emploi : Chez mon papa
| | Sujet: Re: Classification.générale (cours) Mer 5 Déc 2012 - 23:49 | |
| III. EMBRANCHEMENT DES BRYOPHYTESIII.1 CARACTERES GENERAUX1. Vie terricole
: plantes vivant sur le milieu terrestre, c'est-à-dire qui exploitent
le sol par un appareil suceur ( eau est sels minéraux )et épanouissent
dans l'atmosphère un appareil chlorophyllien destiné à produire de la
matière organique.2. Appareil végétatif.
Tiges + feuilles ; pas de véritables racines. Ce sont des poils
rizhoïdes qui remplissent les fonction de pompage nutritif dans le sol
et d'ancrage au sol. Quelques Briophytes sont dépourvues de tiges et de
feuilles : ils ont un thalle et se situent donc en position
intermédiaire entre l'embranchement des Thallophytes ( exemple :
Marchanta polymorpha ) 3. Reproduction
: oogamie . par conséquent grande différence morphologique entre les
organes reproducteurs ( = gamétanges ) mâles et femelles .o. r. mâle : anthéridieso.r. femelle : archégones ou oogones .  4. Les organes reproducteurs sont entourés par une paroi assisiale 4. Les organes reproducteurs sont entourés par une paroi assisiale
( composée d'une ou plusieur assises cellulaires ) et non par une
membrane comme chez les Thallophytes. La paroi assisiale se retrouvera
également dans les Embranchements. 5. Cycle haplo-diplobionte. Grande différence morphologique entre les deux phases, mais c'est la phase haploïde qui domine. Dans
les Embranchements suivants : cycle haplo-diplobionte, mais la phase
diploïde sera dominante. Chez les Thallophytes à caractère
haplo-diplobionte, il y auvait équivalence entre les les phases haploïde
et diploïde .III.2 CLASSES1. Les hépatiques Classe
hétérogène. Thalle appliqué contre le sol, avec rhizoïdes, donc
intermédiaires entre thallophytes et mousses, Exemple déjà cité :
Marchantia polymorpha.2. Les moussesTiges
dressées avec feuilles avec feuilles en vue de l'exploitation de
l'atmosphère. Ceci traduit un perfectionnement dans l'exploitation du
milieu. III.3 LA CLASSE DE MOUSSES.III. 3.1 Développement végétatif des moussesPartons d'une spore jonchant le sol. Elle germe en un filament vert, pluricellulaire et mince appelé protonémaLe protonéma III.3 LA CLASSE DE MOUSSES.III. 3.1 Développement végétatif des moussesPartons d'une spore jonchant le sol. Elle germe en un filament vert, pluricellulaire et mince appelé protonémaLe protonéma
envoie çà et là quelques rhizoïdes dans le sol; il développe à d'autre
endroits des cellules qui deviendront des initiales. Les initiales ont
une forme spéciale ( pyramide à 4 faces ) et se divisent de manière à
produire un méristème puis une tige.Des tiges de
mousses apparaissent ainsi sur le filament et sont groupées plus ou
moins densément ( coussinets ). Ces tiges sont composées de : - une assise externe ou épiderme. - des cellules allongées et étroites sous l'initiale.- du parenchyme vert, turgescent et méatique occupant le reste du volume de la tige.- Tiges et
feuilles sont des structures simples : pas des vaisseaux, pas de tubes
criblés, les mouvements de sèves se font de cellule à cellule.  III.3.2 ReproductionSoit une mousse monoïque ( il existe des mousses dioïques ). En général les archégones ( ou gamétanges femelles ) sont situées au sommet de la tige.Elles présentent en général une forme de bouteille : la base contient l'oosphère ( = gamète femelle ) . L'oosphère est surmontée par des cellules de canal qui occupent l'intérieur du col de la bouteille .Les anthéridies III.3.2 ReproductionSoit une mousse monoïque ( il existe des mousses dioïques ). En général les archégones ( ou gamétanges femelles ) sont situées au sommet de la tige.Elles présentent en général une forme de bouteille : la base contient l'oosphère ( = gamète femelle ) . L'oosphère est surmontée par des cellules de canal qui occupent l'intérieur du col de la bouteille .Les anthéridies
ou gamétange mâles sont situées à l'aisselle des feuilles supérieures.
Elles comprennent comme tout gamétange , une assise de paroi, à
l'intérieur se développent les anthérozoïdes ou gamètes mâles. La fécondation La fécondation
se fait à l'intervention d'une goutte d'eau ( rosée pluie ). Les
anthérozoïdes sont biciliés ; ils nagent à l'intérieur du milieu aqueux
aussitôt après l'ouverture de l'antéridie. Les anthérozoïdes sont
attirés vers le sommet de l'archégone par un phénomène de chimiotactisme
( sécrétion d'acide malique ) - les gamètes mâles ( = anthérozoïdes )
pénètrent à l'intérieur de l'archégone dont les cellules de canal sont
entretemps gélifiées, assurant ainsi une continuité entre la goutte et
l'oosphère.Fécondation de l'oosphère et formation d'un zygote 2n Le zygote se
développe en un corps massif ( qui s'accroît par une cellule initiale
pyramidale e ), à l'intérieur de l'archégone. Le corps massif de garniture 2n évolue en une urne
pourvue d'un pédicelle. Entretemps, l'archégone s'est déchirée et le
sommet de l'urne 2n est coiffé par le sommet de l'archégone ( n ). Les
cellules n qui surplombent l'urne forment la coiffe .  Remarques : 1) Tout ce qui provient du zygote est nécessairement diploïde ( 2n) ; la plante de mousse est, par contre, haploïde.2) L'urne ou sporogone et son pédicelle sont appelé le sporophyte ( 2n ); c'est la partie de la plante qui redonnera des spores.3) Remarques : 1) Tout ce qui provient du zygote est nécessairement diploïde ( 2n) ; la plante de mousse est, par contre, haploïde.2) L'urne ou sporogone et son pédicelle sont appelé le sporophyte ( 2n ); c'est la partie de la plante qui redonnera des spores.3)
La plante de mousse est issue de la spore via le protonéma : sa
garniture chromosomique est haploïde ( n) ; elle est appelée le gamétophyte parce que c'est la partie de la plante qui a produit les gamètes4)
Dans le cas des mousses : le sporophyte ( 2n ) est porté par amétophyte
(n) ou encore la sporophase est tributaire de la gamétophase, car elle
dépend de la phase n qui est le plant proprement dit.Composition de l'urne ou sporogone : coupe longitudinale et transversale. - extérieurement : épiderme - au centre: parenchyme incolore appelé columelle- autour de la columelle : une gaine de grandes cellules appelée archéspores.- entre la columelle et l'épiderme : filaments verts, ramifiés et cloisonnés  SporogénèseChacune des archéspores ( 2n ) subit de nombreuses divisions. L'ensemble des cellules filles issues de chaque archéspore SporogénèseChacune des archéspores ( 2n ) subit de nombreuses divisions. L'ensemble des cellules filles issues de chaque archéspore
constitue un massif sporogonial ( 2n ) qui a un moment donné, se
dissocie et libère un grand nombre de petites cellules appelées sporogonies (2n) . A A
ce moment, les filaments verts de la columelle se transforment ( grâce
à des ferments ) en une bouillie qui sert de nourriture aux
sporogonies.Les sporogonies grossissent et deviennent des sporocytes ( 2n) . Les sporocytes, gorgés de réserves subissent les cinèses réductionnelles : chaque sporocyte donne une tétraspore ( tétra = 4 ) de garniture n.La
tétraspore se disloque ; chaque spore (n) s'arrondit s'arrondit,
s'entoure d'une seconde membrane ( de subérine, plissée) de protection.Lorsque
les spores (n) sont mûres, le sporogone s'ouvre transversalement par
une sorte de couverte toujours surmonté par la coiffe ( n ) qui
constituait le col et le sommet de l'archégone. Les
spores ainsi mises en liberté recommencent le cycle pour autant
qu'elles retombent sur un milieu favorable à leur germination.   JUSTIFICATION DE LA PETITESSE DES MOUSSES1) Pas de racines.2) Pas de tissus JUSTIFICATION DE LA PETITESSE DES MOUSSES1) Pas de racines.2) Pas de tissus
conducteurs; les seuls tissus définitifs d'une mousse sont le
parenchyme et l'épiderme . Donc pas de spécialisation histologique.3) L'urne et son
pédicelle, c'est à dire le sporophyte ( 2n ) est porté par le
gamétophyte ; le sporophyte dépend du gamétophyte pour son alimentation :
il est nécessairement plus petit que le gamétophyte .4) La
fécondation passe obligatoirement par une goutte d'eau: ceci postule un
espace très court entre les gamètes mâles et femelles ; de telles
plantes doivent donc rester petites . Shéma du cycle biologique  GENRE PRINCIPAUX - Distribution Près de 40 espèces en Belgique, avec, pour genres principaux :- Polytrichum- Leucobryum- Eurynehium- Catharinea- Dicranum- DicranellaForment la strate mucinale des prairies et des bois, vivant collées au sol.Sont aussi des plantes épiphytes IV. EMBRANCHEMENT DES PTERIDOPHYTES IV.1 DEJA ACQUIS ( Briophytes )1) Vie terricole2) Paroi des gamétanges de nature assisiale3) Oogamie parfaite4) GENRE PRINCIPAUX - Distribution Près de 40 espèces en Belgique, avec, pour genres principaux :- Polytrichum- Leucobryum- Eurynehium- Catharinea- Dicranum- DicranellaForment la strate mucinale des prairies et des bois, vivant collées au sol.Sont aussi des plantes épiphytes IV. EMBRANCHEMENT DES PTERIDOPHYTES IV.1 DEJA ACQUIS ( Briophytes )1) Vie terricole2) Paroi des gamétanges de nature assisiale3) Oogamie parfaite4)
Haplo-diplophase avec inégalité morphologique accusée entre les deux
phases - chez les Briophytes, la phase (n) est la plus développée ,
c'est-à-dire le gamétophyte - chez les ptéridophytes, le sporophyte ( ou
phase (2n) ) sera la plante proprement dite. IV.2 CARACTERES NOUVEAUX 1) Appareil végétatif complet : racine + tige + feuille.2) Formation d'un embryon.3) Sporophyte de grande taille : c'est la plante proprement dite.4) Différenciation histologique
= conséquence du point précedent. Tissus conducteurs ( vaisseaux
scalariformes ) et de soutien présents : les Ptéridophytes sont appelé Cryptogames vasculaires.5) Hétérosporie : deux types de spores, macro et microspores chez les fougères les plus évoluées.IV.3 CLASSES ACTUELLESIV.3.1 Classification traditionnelleOn
distingue généralement 4 classes ( que certains assimilent parfois à
des Sous-embranchement ou même des Embranchements ) , trois d'etre
elles nous intéressent ;Dans l'ancienne classification en Classes on écrivait il y a quatre classes de fougères que voiçi :1) Les Filicinae = Filicopsides ou classe des fougères proprement dites : Ce sont les fougères ordinaires, dont l'ordre principal est celui des Filicales.2) Sphénopsides = Articulatae ou classe de prèles
: Dont le seul ordre : Equisetales comprend une seule famille (
Equisetacées) et un seul genre ( Equisetum = Prèles ) avec une trentaine
d'Espèces.3) Lycopsides ou classe des Lycopodes : avec deux ordres principaux Lycopodiales et Selaginellales. 4) Psilotopsides : Avec un seul ordre et 3 espèces - pour mémoire.IV.3.2 Nouvelle classification Actuellement on considère des Sous-embranchements équivalents aux anciennes classes :1) Les Ptérophytes constituent de loin le groupe le plus imposant des Ptéridophytes. Ils comprennent deux groupes principaux : les Filicales, auxquelles appartiennent les fougères dont nous avons déjà parlé, sont caractérisées par une isosporie généralisée ; les Hydroptéridales vivent en milieu aquatique et sont hétérosporées. groupe équivalent aux Filicopsides2) Les Sphénophytes sont représentés actuellement par le seul genre Equisetum,
les prêles. Celles-ci développent un rhizome souterrain, muni de
racines et portant des ramifications aériennes annuelles, typiquement
cannelées et pourvues de verticilles de feuilles étroites. Les
sporanges sont groupés en épis à l'extrémité des axes végétatifs ou
d'axes spécialisés, les sporangiophores. Il y a un seul type de spores
(isospories) entourées d'élatères qui contribuent à la dissémination en
se déployant lorsque le taux d'hygrométrie diminue. groupe équivalent aux Sphénopsides3) Les Lycophytes
sont plus diversifiés, avec trois groupes principaux : les lycopodes,
les sélaginelles et les isoètes. Leurs feuilles sont généralement de
petite taille et pourvue d'une seule nervure médiane. - La tige des
lycopodes est rampante, pourvue de racines et porte des axes dressés
(Photo 17) sur lesquels se forment les sporanges dispersés le long de
la tige ou groupés en strobile, sorte d'épi. Les sporanges
sont généralement situés sur la face supérieure des feuilles. Les
méiospores sont toutes de même taille (isosporie).- Les
sélaginelles ont déjà reçu notre attention car c'est au sein de ce
groupe qu'apparaît l'hétérosporie, associée à une hétérosporangie
marquée. -
Les isoètes sont des plantes à tige courte renflée en bulbe. Les
sporanges sont inclus dans la base des feuilles qui, toutes, sont
potentiellement des sporophylles. Il y a hétérosporie. groupe équivalent aux Lycopsides4) Les Psilophytes, dont l'exemple classique est le Psilotum, constituent le groupe le plus primitif des Ptéridophytes. Le Psilotum
est une plante des régions tropicales et subtropicales constituée d'un
rhizome, dépourvu de racine mais nanti de rhizoïdes bi- ou tri-
cellulaires, portant des axes dressés, se ramifiant par dichotomie, sur
lesquels s'insèrent des feuilles ou écailles non vascularisées. Les
sporanges, groupés par trois, sont situés au sommet d'un pédicelle
vascularisé, très court. Les méiospores sont toutes de même taille
(isosporie). groupe équivalent aux PsilotopsidesLe
gamétophyte devient réduit; il sert seulement à produire les gamètes et
à nourrir le zygote jusqu'à ce que l'embryon puisse envoyer une racine
dans le sol.IV.4 CLASSE DES FILICINAE-FILICOPSIDES/ SOUS-EMBRANCHEMENT DES PTEROPHYTES  Le gamétophyte. Schémas.Nous partons d'une spore n qui germe en une petite lame verte ayant une surface horizontale de quelques mm2, c'est le prothalle de la fougèreCe Le gamétophyte. Schémas.Nous partons d'une spore n qui germe en une petite lame verte ayant une surface horizontale de quelques mm2, c'est le prothalle de la fougèreCe
prothalle s'accroît par une initiale; il prend unn aspect cordiforme et
se compose de deux parties uniassisiales séparées par un coussinet
pluricellulaire ou les cellules sont de plus petite taille, le coussinet
envoie dans le sol des poils rhizoïdes.  Le coussinet joue trois rôles :- Il forme des poils rhizoïdes, à rôle nutritif.- Il formera les archégones et les anthéridies.- Il nourrira ultérieurement le zygote; Le prothalle est le gamétophyte de la fougère : il matérialise la phase haploïde qui se termine par la formation des gamètes. Le coussinet joue trois rôles :- Il forme des poils rhizoïdes, à rôle nutritif.- Il formera les archégones et les anthéridies.- Il nourrira ultérieurement le zygote; Le prothalle est le gamétophyte de la fougère : il matérialise la phase haploïde qui se termine par la formation des gamètes.   Au moment de Au moment de
l'apparition des gamètes , la face inférieure de la lame uniassisiale du
prothalle comporte une anthéridie avec un pédicelle . A l'intérieur de
l'anthéridie : des anthérozoïdes ou gamètes mâles pourvus d'une couronne
de cils . Dans les tissus
du coussinet s'est formée une archégone, composée d'une assise
cellulaire de paroi, d'une oosphère ou gamète femelle surmontée de
cellules de canal.La fécondation Lorsque les
Anthérozoïdes sont mûrs, l'anthéridie libère son contenu à l'intérieur
d'une goutte d'eau. Les anthérozoïdes se dirigent vers l'archégone par
réaction de chimiotaxisme ; les cellules de canal se gélifient; il y a
plasmogamie puis caryogamie.Le zygote 2n se
divise 2 fois par cinèses normales donnant par conséquent quatre
cellules filles 2n , appelées les quatre quadrants du Ptéridophyte:
c'est l'embryon.Chaque cellule du quadrant est appelée à un rôle particulier :Le premier quadrant va devenir le pied par prolifération à l'intérieur du prothalle et digérer ce dernier au profil de l'embryonLe second quadrant : va s'enfoncer dans le sol et constituer la racine du Ptéridophyte.Le troisième quadrant est le point de départ de la tige ( Rhizome ) ; il sera peu développé au début.Le quatrième
quadrant deviendra le première feuille et se redressera vers le haut (
les feuilles de fougères sont appelées frondes). Le développement du sporophyteL'embryon va évoluer de manière à devenir la plante de fougère : a) le pied disparaît lorsque la plante sera capable de puiser sa nourriture dans le sol.b) la tige demeure souterraine sauf exception ( rhizome ) et forme un méristème terminal c) les Le développement du sporophyteL'embryon va évoluer de manière à devenir la plante de fougère : a) le pied disparaît lorsque la plante sera capable de puiser sa nourriture dans le sol.b) la tige demeure souterraine sauf exception ( rhizome ) et forme un méristème terminal c) les
feuilles se forment latéralement. Elles se forment latéralement , elles
s'érigent en prenant la forme d'une crosse , d'où leur nom de fronde
avant de s'épanouir. Dans la suite, les feuilles deviennent composées
pennées ou restent simples. Elles sont souvent la seule partie visible
de la fougère sauf en région tropicale.d)
La racine devient pivotante. D'autres racines, adventives ,
apparaissent sur le rhizome. Ainsi se développe la plante de fougère
proprement dite : c'est le sporophyte ( 2n ). Rappel de la mousse: la
plante de mousse était le gamétophyte. Formation des sporanges et sporogénèse. Les sporanges apparaissent à la face inférieure des frondes ; ils sont groupés densément en des ensembles ou sores.Ces sporanges sont généralement protégés par une expansion épidermique appelée indusie. L'indusie
et les sores qu'elle recouvre est située de part et d'autre de la
nervure médiane ou parfois elle est assurée par un rebord de limbe.Les
sporanges naissent à partir d'une cellule épidermique, à la face
inférieure de la fronde ainsi qu'il a déjà été précisé. Une initiale
pyramidale se forme à partir de cette cellule épidemique , et le
sporange formé par le jeu de divisions de l'initiale se compose de :- Un pédicelle.- Une assise de paroi : la plus externe.- Une assise de tapis composée de grandes cellules ( = réserve alimentaire ).- Une cellule centrale : l'initiale qui devient une archéspore.Le
tout est 2n , y compris l'archéspore puisqu'il s'agit du sporophyte.
L'archéspore subit la sporogenèse, déjà décrite chez les mousses , en
suivant le schéma : Archéspores 2n ----> massif
sporogonial ----> sporogonies 2n----> sporocytes
2n ----> méiose R ----> tétraspores n ----> spores n
individualisées en intine et exine .Le sporange 2n adulte renferme de nombreuses spores .Le
sporange s'ouvre et libère es spores. L'ouverture du sporange est
réalisée au moyen d'un anneau de déhiscence. Cet anneau de déhiscence
rappelle par son fonctionnement celui de l'assise mécanique qui préside à
l'ouverture des anthères. Il sera décrit plus loin.  CYCLE BIOLOGIQUE DES FOUGERES CYCLE BIOLOGIQUE DES FOUGERES   Cycle de vie d'une fougère. Les structures haploïdes sont dessinées en rouge. 1 Sporophyte maturea Pétioleb Fronde circinée (= enroulée)c Rhizomed Racines adventices2 Face inférieure d'un segment ou pennea Sore3 Sore en coupea Epiderme supérieurb Tissus spongieuxc Epiderme inférieurd Indusie4 Sporangea Pédonculeb Anneau mécaniquec Cellule de la lèvred Spores5 Jeune prothalle (= gamétophyte)a Rhizoïdesb Paroi de la spore6 Prothalle mature (= gamétophyte)a Rhizoïdes7 Anthéridiea Cellule annulaireb Cellule operculaire8 Anthérozoïdes9 Archégone avec o(v)osphèrea Ventreb Colc Cellule du col10 Zygote dans l'archégone11 Sporophyte embryonnaire12 Sporophyte sur le gamétophyte COMPARAISON ENTRE LE CYCLE DES MOUSSE ET LE CYCLE DES FOUGERESRessemblancesCycle haplo-diplobionte et gamétophyte de petite tailleDissemblancesChez les mousses : le zygote et toutes les cellules qui proviennent de lui sont situées à la partie supérieure du gamétophyte à quelques distances du solLe sporophyte est donc dépendant du gamétophyte pour sa nourriture.Chez les fougères, le sporophyte est au contact du sol :
l'embryon envoie dans le sol une racine qui le rend indépendant du
gamétophyte. C'est donc en raison de la situation du zygote sur le
gamétophyte que les fougères peuvent devenir des plantes de grande
dimension.Le zygote se développe en un embryon
, que l'on rencontre pour la pemière fois et qui est le prélude d'un
sporophyte autonome pouvant acquérir une grande taille et comportant un
appareil élaborant étalé dans l'atmosphère.En outre, il existe un appareil radiculaire véritable qui s'enfonce dans le sol.Un tel sporophyte requiert la spécialisation des tissus formant ses organes d'où une différenciation histologique est tissus de conduction, de soutien et de protection.Les
Filicinées sont aussi les premières plantes vasculaires de l'échelle
taxonomique ; elles ont connu leur apogée à l'époque carbonifère ( voir
géologie )Elle
comprend quelques ordres, dont le plus important est celui des
Filicales, avec une dizaine de familles. L'identification des familles
et des genres est parfois délicate .Quelques
plantes : mas, Dryopteris filix femina , Blechnum spicant,... IV.5 CLASSE DES SPHENOPSYDES-ARTICULATAE/ EMBRANCHEMENT DES SPHENOPHYTESC'est
le groupe des Prèles ou queues de cheval avec l'ordre des Equisetales :
les familles des Equisetacées et le genre Equisetum avec une trentaine
d'Espèces.Exemples : E. palustre , E arvense, E.maximus, E. maximum,...sont considérés généralement comme des plantes adventices.Vestiges d'une flore très répandue au Carbonifère: grand arbres à l'origine de nos gisements houillers.Caractère nouveau : hétéroprothallie Chez les Filicinées : spores identiques et prothalle bisexué mâle et femelle d'où isosporie et isoprothallie.Chez les Prèles : isosporie, mais hétéroprothallie : un prothalle mâle portant des anthéridies et un prothalle femelle portant l'archégone . Afin
de limiter la matière à l'essentiel, cette classe est représentée
ci-après par son cycle biologique, établit par comparaison avec celui
des filicinées . CYCLE BIOLOGIQUE EQUISETINEE  CONES      4) 4)
De nombreuses divisions donnant de nombreuses petites cellules n dont
le noyau s'allonge et le cytoplasme produit deux flagelles: ce sont les
Anthérozoïdes.Rem on peut constater que le prothalle mâle issu de la seconde cellule fille issue de la microspore.Le développement de la mégasporeLa mégaspore est enfermée dans son mégasporange et est toujours entourée d'une paroi épaisse. Elle germe en un prothalle intérieur
, c'est-à-dire que le noyau de départ se divise un grand nombre de fois
et devient un symplaste ; des cloisonnement apparaissent et la
mégaspore devient un tissus composé d'une part de grandes cellules à
l'intérieur desquelles se développe une archégone.En
résumé, la mégaspore se développe en un prothalle femelle
pluricellulaire à l'intérieur duquel une archégone se forme, cette
archégone contenant une oosphère ou gamète femelle . schéma : prothalle femelle de SélaginelleLa schéma : prothalle femelle de SélaginelleLa
fécondation se produit à l'intervention d'une goutte d'eau, permettant
aux anthérozoïdes de cheminer vers le col de l'archégone puis vers
l'oosphère ... le zygote se développe en un embryon (sporophyte). | |
| | | | biologista
Sexe :
Messages : 180
Points : 317
Date de naissance : 07/05/1992
Age : 32
Emploi : Chez mon papa
| | Sujet: Re: Classification.générale (cours) Mer 5 Déc 2012 - 23:55 | |
| CYCLE BIOLOGIQUE CONSEQUENCES DE L'HETEROSPORIE AU POINT DE VUE DE L'EVOLUTIONL'hétérosporie marque un progrès biologique considérable dans l'évolution en ce qui concerne la garantie de la reproduction Chez les CONSEQUENCES DE L'HETEROSPORIE AU POINT DE VUE DE L'EVOLUTIONL'hétérosporie marque un progrès biologique considérable dans l'évolution en ce qui concerne la garantie de la reproduction Chez les
fougères, la plante forme des spores très petites qui doivent gemer en
un prothalle, lequel prothalle devra subvenir par son propre travail
photosynthétique et sa propre alimentation minérale aux besoins de l'embryon. Chez la
Sélaginelle , la plante forme des mégaspores de grande taille qu'elle
bourre de réserves alimentaires, assurant ainsi elle même et à l'avance
la nutrition du futur embryon.La sécurité de
la reproduction se trouve de la sorte considérablement accrue et la
plante peut ne former que relativement peu de mégaspores.D'autre part le
gamétophyte mâle ne devant aboutir qu'à la formation de quelques gamètes
mâles , peut être extrêmement réduit. Il est constitué par une seule
cellule prothalienne portant une seule anthéridie. Il est à
remarquer que la reproduction se fait encore à l'intervention d'une
goutte d'eau, ce qui explique l'existence de gamètes mâles ciliés. Cycle de vie d'une prêle du genre Equisetum. Les structures haploïdes sont dessinées en rouge. Gamétophyte: 1 Sporesa Elatères2 Spores germant3 Prothallea Anthéridieb Rhizoïdes4 Prothallea Archégoneb Rhizoïdes5 Anthéridie6 Anthérozoïde7 Archégonea Anthérozoïdeb O(v)osphère Sporophyte: 8 Jeune sporophyte sur gamétophytea Racine primaire9 Pousse végétativea Verticillesb Ecaille foliairec Rhizome10 Pousse fertilea Strobileb Ecaille folaire11 Portion de strobile avec 3 sporophyllesa Sporangeb Sporangiophore  Cycle de vie d'un lycopode du genre Lycopodium. Les structures haploïdes sont dessinées en rouge. 1 Sporophytea Racineb Rhizomec Tiged Feuillese Strobile2 Tige du sporophytea Epidermeb Cortexc Pericycled Phloèmee Xylèmef Endodermeg Vestige foliaire3 Strobilea Sporangeb Sporophylle4 Sporange5 Sporophyllea Sporange6 Tétrade de spores7 Spore ayant germéa Champignon endophyteb Enveloppe de la spore8 Gamétophytea Anthéridieb Archégonec Rhizoïded Champignon endophyte9 Anthéridie10 Anthérozoïde11 Archégonea O(v)osphère12 Embryona Suspenseurb Piedc Cotylédon13 Jeune sporophytea Pousseb Piedc Racined Gamétophyte  Cycle de vie d'une sélaginelle du genre Selaginella. Les structures haploïdes sont dessinées en rouge. 1 Sporophyte maturea Rhizophoreb Feuillec Strobile2 Strobilea Sporophylleb Ligule3 Microsporangea Tapetumb Cellule-mère de microspore4 Microsporangea Tétrade de microspores5 Microgamétophytea Cellule prothallialeb Cellule d'enveloppec Jeunes anthérozoïdes6 Microgamétophyte maturea Paroi de la microspore7 Anthérozoïde8 Mégasporangea Mégagamétocyteb Tapetum9 Mégasporangea Tétrade de mégaspores10 Mégasporea Mégagamétophyteb O(v)osphèrec Col de l'archégoned Paroi de la mégaspore11 Sporophyte embryonnairea Rhizoïdeb Suspenseurc Rhizophore primaired Pied12 Développement du sporophytea Cotyledonb Tige sous cotylédonsc Rhizophore primaired Paroi de la mégaspore  Cycle de vie de Psilotum. Les structures haploïdes sont dessinées en rouge. 1 Sporophytea Rhizoïdesb Rhizomec Tige aérienned Feuille écailleuse2 Tigea Epidermeb Parenchyme photosynthétiquec Phloèmed Xylèmee Stomate3 Sporange4 Sporange déhiscent5 Spores6 Gamétophytea Archégoneb Anthéridiec Rhizoïded Champignon endophyte7 Anthéridie8 Anthérozoïdes multiciliés9 Archégonea O(v)osphère10 Zygote dans l'archégonea Gamétophyte Illustrations de cycles issues du site : http://www.afblum.be/bioafb/cyclpter/cyclpter.htm Trois
sous-embranchements entièrement fossiles sont habituellement reconnus.
Ceux-ci ont été dominants du milieu du silurien au milieu du dévonien
(425 à 370 millions d'années) à la fin duquel ils ont disparu (environ
360 millions d'années) ; • les Rhyniophytes (425 à 380 millions d'années) • les Zostérophyllophytes (408 à 370 millions d'années)• les Trimérophytes (395 à 375 millions d'années) V. EMBRANCHEMENT DES SPERMATOPHYTES/CORMOPHYTES/PHANEROGAMES V. 1. CARACTERES GENERAUX. 1. Déjà acquisa. Cycle haplo-diplophasique avec prédominance de la sporophase. b. Appareil végétatif complet : tiges + feuilles + racinesc. Mégaspores et microsporesd. Embryon, fût-il rudimentaire2. Caractère nouveau : apparition d'une graineLa graine est un élément complexe de reproduction et de dissémination car elle contient :- Un ébauche de plante complète- D'importantes réserves alimentaires- Une enveloppe protrectrice efficaceDans le sous-embranchement des Gymnospermes
( = graines et ovules nues ), les graines se développent à l'aisselle
d'une écaille étalée ( l'écaille est la bractée du cône ).Dans le
sous-embrauchement des Angiospermes , les graines - et les ovules - sont
enfermés dans un cavité close, l'ovaire ( Aggéion = vase, boîte ;
sperma = graine ).-----------------------------------------------------------------------V. 2. LE SOUS - EMBRANCHEMENT DES GYMNOSPERMES Peu représentées
actuellement à la surface du globe (700 espèces environ), les
Gymnospermes constituent le reliquat d'un ensemble autrefois très
important qui, à son apogée au Jurassique (environ 150 millions
d'années), comptait quelque 20.000 espèces.
Plusieurs caractères différencient les Gymnospermes des Angiospermes :
• leurs ovules nus (avec une certaine tendance à l'angiospermie) ;
• des « fleurs » généralement unisexuées ;
• un sac embryonnaire (gamétophyte femelle) pluricellulaire (7 cellules, le plus souvent, chez les Angiospermes y compris le gamète) ;
• un gamétophyte mâle pluricellulaire (3 cellules chez les Angiospermes, y compris 2 gamètes) ;
• la pollinisation
(phénomène nouveau dans l'évolution des plantes qui désigne la
rencontre du grain de pollen et de la structure femelle) qui se fait
toujours directement sur l'ovule par l'intermédiaire d'une goutte de
pollinisation située dans le micropyle (la goutte micropylaire) et la
germination du pollen qui s'effectue sur le nucelle (alors que chez les
Angiospermes, pollinisation et germination du pollen s'effectuent sur le
stigmate de la fleur) ;
• une surface adaptée de l'enveloppe qui entoure les ovules ;
• une fécondation simple le plus souvent alors qu'elle est double chez les Angiospermes ;
• un tissu de réserve haploïde (triploïde chez les Angiospermes) dans la graine ;
•
des processus de croissance secondaire confèrent, le plus souvent une
forme d'arbre ou d'arbrisseau ligneux au sporophyte des Gymnospermes
mais habituellement, le xylème est composé de trachéides et ne
différencie pas de vaisseaux. 7 Classes , citées par leur ordre d'évolution. pour mémoire. 1) Ptéridospermes : chaînon entre les Ptéridophytes ( frondes ) et spermatophytes ( graines ). Entièrement fossiles.Les Ptéridospermales( 1) Ptéridospermes : chaînon entre les Ptéridophytes ( frondes ) et spermatophytes ( graines ). Entièrement fossiles.Les Ptéridospermales(
fougères à graines ) sont un vaste embranchement de plantes vasculaires
éteintes qui vécurent du Dénovien jusqu'au Jurassique. Elles sont
caractérisées par l'association de frondes filicéennes et d'un appareil
reproducteur sous forme d'ovules, comme chez les plantes supérieures.L'appareil végétatif est apparenté aux fougères tandis que l'appareil reproducteur est comparable à celui des conifères actuels. En En
fait, ces plantes ne portent jamais de graines véritables mais des
ovules et l'état de graine n'est atteint que lorsque l'ovule est séparé
de la plante mère.Le
caractère intermédiaire de ce groupe du paléozoïque joue un rôle
essentiel dans la phylogénie des plantes vasculaires en établissant un
jalon entre les ptéridophytes et les spermatophytes.Les ptéridospermales renferment deux familles, les Lyginopteridacées et les Medullossagées2) Bennettitales : fossiles.3) Cordaïtes : fossiles . Il s'agissait Il s'agissait
d'arbres assez grands avec un port différent de celui des conifères avec
des feuilles rubanées, minces et allongées. Le diamètre du tronc
pouvait atteindre 1,50 m. C'étaient les plus grands arbres du Primaire,
ils affectionnaient les sols humides.Les cônes
étaient moins complexes que chez les conifères actuels. Les fleurs mâles
étaient formées d'étamines à plusieurs sacs polliniques et les fleurs
femelles d'ovules disposés en cône. Il y a toujours zoïdogamie et la
graine se détache à l'état non embryonné4) Cycadales
: La plupart fossiles, mais il reste encore quelques représentants de
cette classe dans notre flore actuelle : Cycas, Encophalartos. 5) Ginkgoales, avec un représentant encore actuel : Ginkgo biloba, espèce dioïque. 5) Ginkgoales, avec un représentant encore actuel : Ginkgo biloba, espèce dioïque.  Ils datent du primaire et une seule espèce subsiste. Elle se caractérise par les feuilles à limbe en éventail. Le ginkgo bilobaest la seule espèce qui a survécu jusqu'à ce jour. Il se retrouve dans les régions montagneuses de la Chine.Les Ils datent du primaire et une seule espèce subsiste. Elle se caractérise par les feuilles à limbe en éventail. Le ginkgo bilobaest la seule espèce qui a survécu jusqu'à ce jour. Il se retrouve dans les régions montagneuses de la Chine.Les
feuilles cunéiformes de 4 à 6 centimètres, sont en éventail divisé en
deux lobes et souvent groupées par 3 ou 5 sur de courts dards. Vert
tendre en été, elles tournent au jaune or en automne. L'arbre peut
atteindre 20 mètres de hauteur par 13 mètres de largeur. Les fleurs
mâles en chaton isolés, ont des étamines à deux sacs polliniques. Les
fleurs femelles, longuement pédonculées, sont formées de deux carpelles,
à ovule pourvu d'une chambre pollinique; il y a zoïdogamie. Le fruit
est une drupe à amande comestible de 2,5 centimètre. Selon le climat, la
graine est embryonnée ou non quand elle se détache de l'arbre. Au printemps I, les Ginkgo mâles libèrent les grains de pollen. Les Ginkgo
femelles produisent des ovules groupés par deux au sommet de
pédicelles. La chambre sous-micropylaire se remplit d'un liquide
visqueux qui perle par le micropyle. Le pollen est capturé par une
gouttelette visqueuse. Le liquide se rétracte dans la chambre sous
micropylaire et le micropyle se referme. Dans le liquide, le pollen
germe, ce qui déclenche l'accroissement de l'ovule et l'accumulation de
réserves. En fin d'été, les ovules mûrissent et
deviennent semblable à des prunes jaunes. Dans la chambre
sous-micropylaire, le protalle mâle issu du pollen, évolue lentement. En automne,
après la chute des feuilles, les ovules (drupes) tombent au sol. La
partie externe du tégument, devenue une pulpe jaune se décompose. Les
fausses prunes du Ginkgo ne sont pas des fruits ni une graine, c'est un oeuf. En début d'hiver,
le prothalle mâle produit des spermatozoïdes flagellés qui vont
féconder l'oosphère. Dès la fécondation, l'embryon commence son
développement. Au printemps II, l'embryon poursuit sa croissance, sort de l'ovule, il germe pour produire une plante autonome.Le
Ginkgo est souvent planté dans nos ville car il résiste bien à la
pollution. Les horticulteurs ne plantent que des sujets mâles, car les
femelles produisent des fruits nauséabonds. En Chine on préfère les
femelles pour les amandes très nutritives.6) Coniférales:
cette classe s'identifie actuellement avec le sous-embranchement des
Gymnospermes : ce sont les "conifères " ou " résineux " du langage
courant. Représentée par environ 300 espèces. 7) Gnétales : deux genres actuels : Gnetum et Welwitschia. Soudure avec les Angiosperme par le gamétophyte femelle.  V.3 LA CLASSE DES CONIFERES V.3.1. De manière résumée : 3 palliers évolutifs importants au sein des Gymnospermes 1) La classe des Cycadales V.3 LA CLASSE DES CONIFERES V.3.1. De manière résumée : 3 palliers évolutifs importants au sein des Gymnospermes 1) La classe des Cycadales
est encore pourvue d'anthérozoïdes ciliés ( en forme de toupie ) et le
prothalle femelle est un tissus pluricellulaire avec archégones ( Les
Gynkoales ont un cycle voisin de celui des Cycadales ).2) Chez les Coniférales,
il y a disparition des gamètes mâles ciliés , et apparition d'un boyau
pollinique ( = siphonogamie ). Le prothalle femelle est encore
pluricellullaire avec archégones.3) Chez les Gnéthales : l'évolution vers les Angiospermes se situe au niveau du prothalle femelle . - Chez Welwitschia: prothalle femelle articulaire et dépourvu d'archégones .- Le Gnétum:
prothalle femelle symplastique avec un nombre indéterminé de noyaux . Ce
prothalle femelle constitue une transition vers les Angiospermes qui
possèdent un prothalle femelle symplastique avec huit noyaux.---------------------------------------------------------------------------------------------------------------- Ce bref survol
de l'évolution chez les Gymnospermes, en deça et au delà de la classe
des Coniférales nous permet de mieux comprendre le cycle biologique de
nos "résineux" actuels, étant entendu qu'ils constituent eux mêmes une
"étape" vers le cycle biologique des Angiospermes. V.3.2 Le cycle biologique des Coniférales : description. V.3.2.1 Le sporophyte des ConiféralesArbres ou
arbustes à aiguilles ou à écailles le plus souvent persistantes pendant
plusieurs années ; généralement monoïques ( parfois dioïques ) . Epicéa, pin, sapin, if, mélèze, thuya,... sont qualifiés de résineux en Sylviculture.     Le sporophyte est 2n, il porte des cônes mâle et des cônes femelle, également 2n.Le cône mâle rappelle le cône des Equisétales. Le sporophyte est 2n, il porte des cônes mâle et des cônes femelle, également 2n.Le cône mâle rappelle le cône des Equisétales.  Ce cône est formé par des bractées qui se retroussent de manière à constituer une enveloppe pour les microsporanges situés à la face inférieure des bractées. Les microsporanges sont appelés anthères et contiennent des microspores ou futurs grains de pollen en quantité considérable. Ce cône est formé par des bractées qui se retroussent de manière à constituer une enveloppe pour les microsporanges situés à la face inférieure des bractées. Les microsporanges sont appelés anthères et contiennent des microspores ou futurs grains de pollen en quantité considérable.   Le cône femelle deviendra le strobile mûr, avec grainesAu départ, le Le cône femelle deviendra le strobile mûr, avec grainesAu départ, le
cône femelle se compose d'un naxe porteur d'écailles ovulifères : ces
écailles sont soutenues par une bractée. A la face supérieure de
l'écaille se trouvent deux ovules .Les ovules sont homologues d'un mégasporange ( voir sélaginelle)Description d'un ovule : - au centre : un corps massif ou nucelle- autour du nucelle : un tégument ou enveloppe de l'ovule- l'ouverture du tégument s'appelle micropyle - à l'intérieur du nucelle, une cellule va devenir une archéspore L'archéspore (
2n ) subit la mégasporogénèse ( Archéspore ( 2n ) ---->
Mégasporocyte( 2n )----> méïose------>une seule Mégaspore n ). La mégaspore est
à ce moment la seule celule ( n ) à l'intérieur de l'ovule. La
mégaspore ( n ) divise son noyau et devient un symplaste ( n ) qui se
nourrit partiellement du nucelle ( 2n )Le symplaste se cloisonne et devient un tissus : c'est le prothalle femelle ou endosperme des conifèresA l'intérieur de l'endosperme, vers le micropyle, une cellule se divise et devient une archégone. L'archégone se compose de deux cellules formant le col et d'une cellule sous-jacente : l'oosphère ou gamète femelle . La fécondationLa fécondation La fécondationLa fécondation
est précédée par l'ouverture des écailles ovulifères afin de permettre
la pénétration des grains de pollen; Ceux-çi atteignent les ovules ,
pénètrent à l'intérieur de ceux-çi par le micropyle et aboutissent à la
surface du nucelle . La microspore deviendra grain de pollen après sa chute sur le nucelle où son noyau se divisera un certain nombre de fois : l'issue de ces divisions sera :1) Une grande cellule ou cellule végétative 2) Une petite cellule ou cellule générative3) un tissus de petites cellules Toutes ces cellules sont évidemment de garniture haploïde.  La grande cellule et le tissus des petites cellules forment le prothalle mâle ou gamétophyte mâleLa grande cellule prothalienne ou cellule végétative développe un boyau pollinique La grande cellule et le tissus des petites cellules forment le prothalle mâle ou gamétophyte mâleLa grande cellule prothalienne ou cellule végétative développe un boyau pollinique
qui pénètre à l'intérieur du nucelle, tandis que la cellule générative
prend une cloison transversale qui donnera une cellule de pied et une
cellule de corps.La cellule de corps se détache de la cellule de pied et s'engage à l'intérieur du boyau pollinique . Le boyau Le boyau
pollinique a traversé le nucelle et arrive au contact avec l'archégone. A
ce moment, le noyau de la grande cellule prothalienne dégénère et
disparaît, tandis que le noyau de la cellule de corps subit une division
( qui ne s'explique pas car elle n'est pas nécessaire ; cette division
annonce toutefois la fécondation des Angiospermes ... voir plus loin ).
L'issue de cette caryocinèse est la formation de deux gamètes mâle . Un Un
des gamètes mâle assure la caryogamie avec l'oosphère. Le zygote se
développe en un embryon. L'embryon se développe à l'intérieur d'une
graine . La graine germera en un nouveau sporophyte. Les cotylédons d'une graine de coniférale sont plus ou moins nombreux ( de 2 à 15 ), la germination est épigée.Le tégument de l'ovule s'épaissit, s'imperméabilise ( subérine ), et devient l'enveloppe ou spermoderme.Composition d'une graine de Coniférale 1) Spermoderme : Ancien tégument ( 2n femelle ) de l'ovule ; rôle protecteur.2) Nucelle : ce qu'il en reste ( 2n femelle ) ; rôle nourricier depuis le stade archéspore.3) Endosperme ou prothalle femelle : ( n femelle ) ; rôle nourricier.4) Embryon Les cotylédons d'une graine de coniférale sont plus ou moins nombreux ( de 2 à 15 ), la germination est épigée.Le tégument de l'ovule s'épaissit, s'imperméabilise ( subérine ), et devient l'enveloppe ou spermoderme.Composition d'une graine de Coniférale 1) Spermoderme : Ancien tégument ( 2n femelle ) de l'ovule ; rôle protecteur.2) Nucelle : ce qu'il en reste ( 2n femelle ) ; rôle nourricier depuis le stade archéspore.3) Endosperme ou prothalle femelle : ( n femelle ) ; rôle nourricier.4) Embryon
reproducteurde la plante ( 2n fécondé ) . L'embryon trouve ses réserves
alimentaires dans l'endosperme puis dans le reste du nucelle.   | |
| | | | biologista
Sexe :
Messages : 180
Points : 317
Date de naissance : 07/05/1992
Age : 32
Emploi : Chez mon papa
| | Sujet: Re: Classification.générale (cours) Mer 5 Déc 2012 - 23:59 | |
| V.4. LE SOUS-EMBRANCHEMENT DES ANGIOSPERMESC'est le groupe
le plus important, quantitativement, parmi les plantes terrestres ( plus
de 300.000 espèces ) et plus particulièrement en agronomie.*Rappelplante monoïque
: Se dit d’une espèce dont les fleurs unisexuées mâles (à étamines) et
femelles (à pistil) sont portées par le même pied. Cela permet la
fécondation croisée (allogamie).
- Le maïs est une plante monoïque.
plante dioïque
:Se dit d'une espèce dont les fleurs unisexuées mâles (à étamines) et
femelles (à pistil) sont portées par des pieds différents.
- Le palmier-dattier ou pommier des landes sont des plantes dioïques.
plante hermaphrodite
: possèdent des fleurs avec des pièces mâles et des pièces femelles
(chaque fleur possède des organes mâles et des organes femelles).
- Le tournesol, la rose, la primevère sont des plantes hermaphrodites, .
V.4.1. Caractères nouveauxa) Ovules enfermés à l'intérieur d'une paroi : la paroi de l'ovaire.b) Les organes reproducteurs sont réunis sous la forme d'une fleur.Le sporophyte : est repésenté par toutes les plantes qui nous sont familières : renoncule, chêne, betteraveLa fleur. Se compose de deux types d'organes :- Reproducteurs : étamines ( anthères ) et pistil ou gynécée ( carpelles ) avec ovules.- Protecteurs :
pétales ( corolle ), sépales ( calice ), bractées. Corolle et calice
constituent le périanthe ( les tépales constituant le périgone c'est à
dire un périanthe homogène ).Schéma d'une fleur complète  Schémas d'un carpelle, d'une étamine ( anthère ) Schémas d'un carpelle, d'une étamine ( anthère ) L'ovaire est la partie renflée du carpelle ; le style ou col est surmonté par le stigmateV.4.2 Développement d'une anthère et formation des grains de pollenAu départ, un anthère est composée de 4 massifs ( les futures loges polliniques ) comportant chacun au centre des archéspores L'ovaire est la partie renflée du carpelle ; le style ou col est surmonté par le stigmateV.4.2 Développement d'une anthère et formation des grains de pollenAu départ, un anthère est composée de 4 massifs ( les futures loges polliniques ) comportant chacun au centre des archéspores
. Les archéspores sont encadrées par une assise de tapis et deux assise
intermédiaires à rôle nourricier . Enfin, un assise " mécanique " à
voir ci après et l'épiderme encadrent chaque loge pollinique en
formation. Les archéspores subissent la microsporogénèse, dont l'issue est la production de microspores (n) avec intine et exine Les archéspores subissent la microsporogénèse, dont l'issue est la production de microspores (n) avec intine et exine archéspores ( 2n) --------> massif sporogonial------->sporogonies ------->sporocytes-------> méiose------->tétrade 4x (n)-------->4 microspores n A l'issue de la microsporogénèse , les anthères se présentent comme suit :  Il se produit Il se produit
une inversion de courbure au nieau des sillons latéraux , grâce a
l'inversion de la forme trapézoïdale des cellules appartenant à l'assise
mécanique.Le mouvement
d'inversion se pousuit et aboutit à la déchirure des loges polliniques ,
qui entretemps ont conflué deux à deux, de part et d'autre d'un sillon
médian. Libération des spores.  REMARQUES1) REMARQUES1)
Il arrive que la déhiscence des anthères soit transversale ou encore
que l'ouverture soit réalisée grâce à des pores terminaux .2) Certaines anthères n'ont que deux loges ; d'autres en on plus de quatre.3) Les
microspores sont des cellules uninucléées, de garniture n ; elles
deviendront grains de pollen au moment où les loges polliniques
s'ouvrent ; le grain de pollen est composé de deux cellules : - une grande cellule ou cellule végétative ou prothalle mâle - une petite cellule ou cellule générativeV.4.3. Développement d'un ovule et mégasporogénèseL'ovule est produit par une prolifération locale du placenta : un massif cellulaire se soulève d'abord pour former le nucelle (Figure 28.1) ; ensuite par des divisions périclines, deux bourrelets circulaires, enveloppants, sont produits : ce sont les téguments
(T1 et T2). Chez certains groupes d'Angiospermes, un seul tégument est
formé. Les téguments grandissent en couvrant progressivement le nucelle
mais en laissant libre un pore donnant accès au nucelle, le micropyle. L'ovule ayant atteint sa taille maximale est fixé au placenta par l'intermédiaire d'un petit pied, le funicule. Téguments et nucelle sont soudés à la base.Les ovules sont des mégasporanges ; ils évolueront normalement en graines, tandis que les carpelles donneront des fruits.Le carpelle le
plus simple se compose d'une cavité emplie d'ovules, et se termine par
un stigmate ( exemple de la fleur de perce-neige). La disposition et le nombre d'ovules varient considérablement; il existe des carpelles uniovulés ( Graminées ).  Schéma général d'un ovule d'Angiosperme - 2 téguments ( 1 tégument chez les Coniférales )- nucelle : partie interne de l'ovule- micropyle : ouverture de l'ovule entre les téguments- chalaze = base de l'ovule où s'attache le funicule- funicule = partie inférieure de l'ovule - placenta point d'attache sur l'ovule Il existe des Schéma général d'un ovule d'Angiosperme - 2 téguments ( 1 tégument chez les Coniférales )- nucelle : partie interne de l'ovule- micropyle : ouverture de l'ovule entre les téguments- chalaze = base de l'ovule où s'attache le funicule- funicule = partie inférieure de l'ovule - placenta point d'attache sur l'ovule Il existe des
ovules unitégumentéschez certaine s familles dicotylées : Bétlacées,
Salicacées, Myricacées, Juglandacées, Lauracées, Cornacées,
Ombellifères, certaines renonculacées.D'autres familles peuvent avoir un troisième tégument. Position de l'ovule.1. L'ovule est dit droit ou orthotrope lorsque le micropyle est tourné vers le haut ( funicule et micropyle sur un même axe) ; cas rare.2. Lovule est dit couché ou campylotrope lorsque le micropyle est orienté vers l'horizontale ( angle droit).3. L'ovule est dit renversé ou anatrope lorsque le micropyle est tourné vers la base de l'ovule. C'est le cas le plus fréquent. Position de l'ovule.1. L'ovule est dit droit ou orthotrope lorsque le micropyle est tourné vers le haut ( funicule et micropyle sur un même axe) ; cas rare.2. Lovule est dit couché ou campylotrope lorsque le micropyle est orienté vers l'horizontale ( angle droit).3. L'ovule est dit renversé ou anatrope lorsque le micropyle est tourné vers la base de l'ovule. C'est le cas le plus fréquent.  Mégasporogénèse Mégasporogénèse  Considérons Considérons
un ovule droit : le nucelle et le mycropyle sont tournés vers le haut.
L'ovule appartient au sporophyte et est donc 2n Au sommet du nucelle, sous son épiderme une cellule fonctionne en archéspore,
le stade sporogonial saute . L'archéspore se nourrit de cellules
nucellaires voisines te devient un mégasporocyte ( 2n ). Celui-çi subit
les cinèses réductionnelles,d'où quatres cellules n formant une
tétraspore, cependant trois cellules de la tétraspore sont digérée par
la quatrième : cette drnière devient une mégaspore (n) bourrée de
réserves.Développement du prothalle femelleLa mégaspore se divise à trois reprise et devient un symplaste à 8 noyaux n. ou prothalle femelle Le prothalle
femelle est aussi appelé sac embryonnaire, car c'est à cet endroit que
sera suspendu l'embryon ; il correspond à l'endosperme des Gymnospermes .Les huit noyaux symplastiques portent des noms consacrés : partant du micropyle vient l'oosphère ou gamète femelle ( n ) flanquée de part et d'autre de deux synergides (n) . Vers le centre du symplaste : deux noyaux polaires (n) ; du côté opposé à l'oosphère et aux synergides viennent se placer les trois noyaux antipodes (n) Il est à Il est à
remarquer que beaucoup de plantes ont une gamétogénèse femelle
raccourcie : les quatres noyaux issus de la tétraspore subissent chacun
une cinèse normale et donnent une garniture symplastique à 8 noyaux n. (
exemple : le Lis). | |
| | | | biologista
Sexe :
Messages : 180
Points : 317
Date de naissance : 07/05/1992
Age : 32
Emploi : Chez mon papa
| | Sujet: Re: Classification.générale (cours) Jeu 6 Déc 2012 - 0:03 | |
|  LE BOYAU POLLINIQUE ET LA DOUBLE FECONDATIONRappel :1) Distinction LE BOYAU POLLINIQUE ET LA DOUBLE FECONDATIONRappel :1) Distinction
entre microspore ( 1 cellule n ) et grain de pollen ( 2 cellules:
cellule générative et cellule végétative ou prothalle mâle ) ; le grain
de pollen existe au moment du fonctionnement de l'assise mécanique .2) devenirs- de l'ovule après fécondation = la graine- de l'ovaire après fécondation = le fruit ( sauf cas particulier ).Prenons le cas
simple d'un grain de pollen projeté dans l'atmosphère et qui aboutit sur
le stigmate d'un carpelle à féconder. Plusieurs circonstances
favorisent la fixation du grain de pollen sur les stigmates :- les aspérités de l'exine, augmentant la surface de contact;- les nombreuses cavités existant entre les papilles stigmatiques ; les grains de pollen s'y enfoncent par gravité ;- la viscosité des papilles stigmatiques et des intervalles entre celles-ci : adhérence des grains de pollen renforcée;- la plus ou moins grande surface portante des stigmates, au sommet du col carpellaire.Soit un grain de
pollen immobilisé sur une surface stigmatique . La surface stigmatique
cède de l'eau au grain de pollen ( phénomène d'osmose ) qui gonfle et développe un boyau pollinique à partir de la cellule végétative. Le boyau pollinique pénètre à l'intérieur du col carpellaire, traverse l'ovaire et aboutit au micropyle.  Il Il
est à noter que la longueur du boyau peut être importante , plus
particulièrement dans le cas d'ovules renversés ( ovules anatropes ).Le boyau
pollinique aboutit contre le sac embryonnaire ; à ce moment le noyau
végétatif ou noyau prothallien mâle disparaît, son rôle étant terminé.Le noyau de la
cellule générative s'est engagé dans le boyau pollinique dès que
celui-çi s'est développé à l'intérieur du corps carpellaire , et le
noyau génératif suit à distance le noyau prothallien mâle jusqu'au
moment de sa disparition.Arivé en fin de
parcours du boyau pollinique, le noyau génératif subit une cinèse qui
donne deux gamètes mâles ( ces gamètes ne sont plus ciliés puisqu'il y a
siphonogamie , ils ne portent plus le nom d'anthérozoïdes.  Les membranes de Les membranes de
contact entre le boyau pollinique et la partie femelle se dissolvent à
l'intervention de ferments et les deux gamètes mâles pénètrent à
l'intérieur du prothalle.-l'un des gamètes mâles fusionne avec l'oosphère , ce qui donne un embryon 2n-l'autre gamète
mâle réalise la fusion nucléaire avec les deux noyaux polaires , d'ou
apparition d'un noyau 3n : 2 x femelle et une fois mâle.Le
fait nouveau réside dans l'utilisation des deux gamètes mâles issus du
noyau génératif du grain de pollen , c'est pourquoi on parle de double
fécondation , étant entendu qu'il n'existe qu'un seul zygote. (Chez
les Coniférales, un seul gamète mâle était utilisé =fécondation
simple).  Après Après
plasmogamie et formation des noyaux 2n et 3 n, les autres noyaux de
l'ancien prothalle femelle, c'est-à-dire les 3 antipodes et les deux
synergides , se tire-bouchonnent et disparaissent. REMARQUES-
Nous avons suppposé un cas simplifié à l'extrême de la double
fécondation. plusieurs grains de pollen tombent simultanément sur les
stigmates ; plusieurs ovules sont à féconder dans un ou plusieurs
ovaires.-
Le transport des grains de pollen est réalisé par le vent (
anémophilie ) mais il peut aussi se faire par les insectes ( entomogamie
. Il va de soi que de très nombreux grains de pollen doivent être
produits ( = explication de la microsporogenèse ). -
La maturité des grains de pollen doit correspondre avec celle des
carpelles ( ovules ). Ce n'est pas toujours le cas :protérandrie,
protérogynie.- Il y a fécondation directe ou autogamie lorsque le pollen d'une fleur tombe sur les stigmates de la même fleur ( ou plante ) et la féconde Cas du froment;- Il y a fécondation croisée ou allogamie lorsqu'une fleur est fécondée par un pollen étranger . Seigle, plantes fruitières , arbres forestiers en général.Ces
diverses considérations revêtent leur importance en production
intensive ( plantes pollinisatrices : exemples vergers ) t surtout en
améliorations des plantes Cycle biologique complet des Angiospermes   Caractéristiques importantes du cycle biologique des Angiospermes1. La vraie nature de la fleur (1) Caractéristiques importantes du cycle biologique des Angiospermes1. La vraie nature de la fleur (1)
La fleur est essentiellement un organe qui forme des sporanges et, par
là, des spores, c'est-à-dire des cellules reproductrices agames. Il est
donc abusif de considérer la fleur comme étant un organe de reproduction
sexuée ou même de définir des fleurs comme mâles ou femelles. La mise à
fleur n'est certainement pas un acte de sexualisation. Parmi les «
pièces florales », les unes sont des sporophylles : telles sont les
étamines (microsporophylles) et le carpelle (mégasporophylle). Les
autres forment le périanthe, c'est-à-dire un ensemble de pièces
protectrices. Les sacs polliniques sont donc des microsporanges et le
nucelle, un mégasporange ; autrement dit, l'ovule est un mégasporange
tégumenté. (2) Si la fleur est, au fondement, un organe
de reproduction asexuée, il est évident que la désigner comme un organe
sexué peut se justifier de diverses manières : - Les gamétophytes qui
constituent les véritables organes sexués sont extrêmement réduits et se
développent totalement ou partiellement au sein des organes de la
fleur. Ainsi, la mégaspore – dans laquelle reste entièrement et
définitivement enfermé le gamétophyte femelle – n'est jamais destinée à
être libérée et toute la reproduction sexuée se fait donc au sein du
mégasporange et de la fleur elle-même. - La fleur est finalement
organisée pour conduire à bon terme l'embryon résultant de la
fécondation, qui est le plus sûr critère de la sexualité : c'est donc
avec ces réserves que l'on peut admettre la « sexualisation » de la
fleur. (3) Ainsi définie dans ses fonctions, la fleur
peut-elle être tenue pour l'homologue d'un strobile de Gymnosperme ou
d'une Sélaginelle ? Si la fleur se compose de véritables sporophylles
enveloppées de pièces protectrices, la réponse doit être affirmative.
Si, par contre, la fleur est issue du fonctionnement d'un méristème
distinct de celui qui engendre les primordiums foliaires et gemmaires,
alors il n'y a pas de communauté d'origine entre carpelles ou étamines,
d'une part, et feuilles, d'autre part. Dans ce cas, les premiers ne
seraient pas de vraies sporophylles. Les études génétiques et
moléculaires démontrent que feuilles et organes floraux sont homologues.
La fleur peut donc être considérée comme étant l'homologue du strobile
des Gymnospermes ou de la Sélaginelle. 2. La réduction du gamétophyte (4) L'édification
du gamétophyte femelle implique 3 vagues de mitoses « somatiques »
successives (cas habituel du sac embryonnaire monosporique de type Polygonum).
Un tel sac embryonnaire réduit à 8 noyaux est souvent interprété comme
étant homologue des deux seuls archégones à 4 noyaux que l'on a
notamment rencontré chez les Gymnospermes. Un seul de ces « archégones
», du côté micropylaire, fournirait l'unique gamète femelle (l'oosphère)
accompagné des deux cellules de col (les synergides). L'archégone
opposé serait devenu stérile. Mais cette extrême réduction peut, en
quelques cas, aller plus loin encore. Le noyau mégasporal réduit peut ne
subir que deux mitoses (type Oenothera) ne livrant donc qu'un
seul archégone à 4 noyaux. Ou bien encore, et c'est la réduction maximum
possible dans l'alternance de phases, les 4 noyaux de la tétrade
mégasporale restés libres persistent, ne subissent qu'une seule mitose
et s'organisent en un sac embryonnaire à 8 noyaux (type Adoxa).
L'haplophase est ici réduite à une seule division "somatique" !
L'archéspore devient directement le sac embryonnaire et le stade de
tétrade lui-même est court-circuité (sac embryonnaire tétrasporique) ! (5)
Chez le gamétophyte mâle, le prothalle n'est plus représenté que par
une seule cellule, de même que l'anthéridie. Celle-ci ne subit plus
qu'une seule division pour donner les deux gamètes. Au total,
l'haplophase ne comporte donc ici que deux mitoses somatiques. 3. La nature de la graine L'albumen (6)
La double fécondation constitue un fait marquant de la reproduction
sexuée chez les Angiospermes. Il en résulte la formation, au sein de la
jeune graine, de deux zygotes qui donneront, l'un, l'embryon diploïde de
la plante en devenir et l'autre l'albumen triploïde dont le
cloisonnement ne fournira qu'un massif nourricier et temporaire. Sous ce
rapport, l'albumen et l'endosperme des Gymnospermes sont analogues.
L'analogie s'arrête cependant là car le second correspond, en fait, à la
partie végétative du gamétophyte femelle. La graine (7) La
structure caryologique de la graine est, chez les Angiospermes,
différente de ce qu'elle est chez les Gymnospermes. En effet, ce qui
restait du gamétophyte a été rapidement résorbé par la croissance de
l'embryon et de l'albumen. Dans une graine mûre d'Angiosperme, il n'y a
donc plus de trace du gamétophyte femelle. Une génération est en
quelque sorte « sautée » dans l'alternance de phases : c'est la
génération sporophytique grand-maternelle qui « élève »le jeune
sporophyte devenu orphelin peu après sa naissance. (8)
Mais ce qui est commun au fondement entre la graine des Gymnospermes et
des Angiospermes va permettre de préciser la vraie nature de cet organe
au niveau des Spermatophytes. Chez les Sélaginelles, la mégaspore est
encore une cellule libre, à paroi d'ailleurs très épaisse, tôt ou tard
vouée à la dissémination avec le gamétophyte qu'elle renferme. Rien de
tel dans la vraie graine. Le sac embryonnaire fait partie du nucelle :
il n'est qu'un élément d'un tissu et a perdu toute possibilité de
dispersion. Il est inexorablement destiné à se développer dans l'ovule
et, par là, dans l'ovaire qui l'abrite le cas échéant. Et c'est ce trait
qui conduit la graine à sa véritable destinée et qui manifeste un
tournant décisif de l'évolution végétale propre aux authentiques
Spermatophytes. Alors que chez les Gymnospermes, on retrouve encore, çà
et là, une trace de cette vocation originelle (dissémination) par
l'acquisition d'un certain épaississement de la paroi mégasporale, ce
caractère a complètement disparu chez les Angiospermes. 4. Le fruit (9)
Toutes les dispositions envisagées ci-dessus aboutissent finalement à
une très grande sécurité de la reproduction ; l'embryon est de mieux en
mieux protégé et la fermeture des carpelles, aboutissant au fruit,
ajoute encore à cette protection et la complète. Mais la conséquence en
est que le grain de pollen ne peut plus atteindre directement le
nucelle, comme c'était le cas chez les Gymnospermes. L'efficacité de la
reproduction est cependant assurée car les gamètes mâles sont dorénavant
véhiculés jusqu'à l'ovule par le canal du tube que produit le grain de
pollen après avoir germé sur le stigmate. La protection des gamètes
mâles est, elle aussi, assurée avec grande sécurité. | |
| | | | Contenu sponsorisé
| | Sujet: Re: Classification.générale (cours) | |
| |
| | | | | | Classification.générale (cours) | |
|
Sujets similaires | |
|
| | Permission de ce forum: | Vous ne pouvez pas répondre aux sujets dans ce forum
| |
| |
| | Site partenaire |  |
|